
Box 4
Folder 25. Whales Fossil
Item 5. Ankle Morphology of the Earliest Cetaceans

![]()






Transcribed Text (OCR)
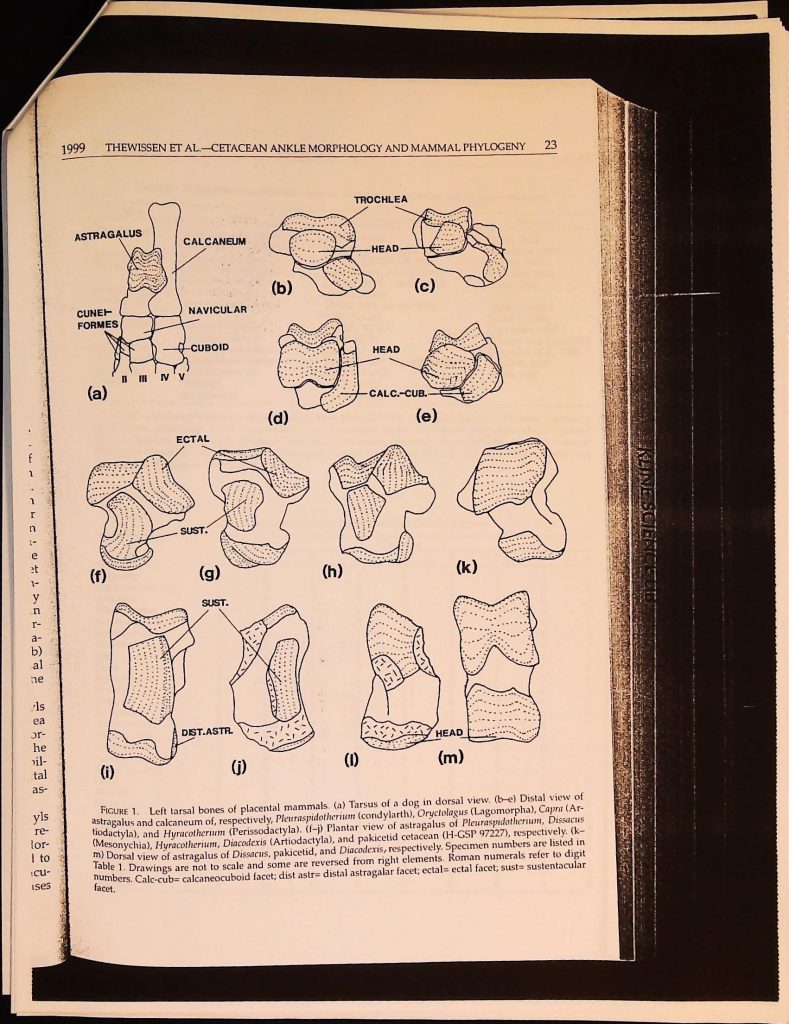
GARY MANGIACOPA ARCHIVE ============================================================ Title: B4F25I5 Slug: b4f25i5 Categories: Cryptozoology Source: https://garymangiacopraarchive.com/b4f25i5 Pages: 10 scanned, 10 extracted OCR: Google Vision API (document_text_detection) Processed: 2026-06-06 ============================================================ Syst. Biol. 48(1):21-30, 1999 Ankle Morphology of the Earliest Cetaceans and Its Implications for the Phylogenetic Relations among Ungulates J. G. M. THEWISSEN' AND S. I. MADAR² 'Department of Anatomy, Northeastern Ohio Universities College of Medicine, Rootstown, Ohio 44240, USA; E-mail: thewisse@neoucom.edu Department of Biology, Hiram College, Hiram, Ohio 44234, USA Abstract. Recent molecular studies are inconsistent with ungulate phylogenetic trees that are based on morphological traits. These inconsistencies especially relate to the position of cetaceans and perissodactyls. Evaluation of the close phylogenetic ties between artiodactyls and cetaceans has been hampered by the absence of tarsal bones of primitive cetaceans, as artiodactyls are often diagnosed on the basis of their tarsus. We here describe newly discovered tarsal bones that are the oldest cetacean tarsals known. We present a character analysis for primitive ungulate tarsals and evaluate their impact on the ungulate phylogenetic tree. Tarsal data are consistent with some molecular studies in suggesting that the extant sister group of Cetacea is Artiodactyla or that Cetacea should be included within the latter order. Tarsal data do not support Cete (Mesonychia plus Cetacea) and are consistent with the exclusion of perissodactyls from paenungulates as suggested by some molecular studies. [Artiodactyla; Cetacea; locomotion; Mesonychia; Ungulata.] The ankle (tarsus) plays a crucial role in the morphological characterization of a number of higher-level clades of mammals. One of the main characters defining Pro- boscidea, for instance, is the presence of a medial process on the astragalus (Tassy, 1996). Archonta (Primates, Dermoptera, Chiroptera, and Scandentia) are charac- terized by fused sustentacular and nav- icular facets on their astragalus (Szalay and Drawhorn, 1980; for an explanation of anatomical terms, see Appendix 1) and all Artiodactyla bear a trochleated astragalar head (Schaeffer, 1947). Despite major mor- phological differences in ankle morphology at higher taxonomic levels (e.g., Matthew, 1937), only minor differences commonly occur within mammalian orders (e.g., ar- tiodactyls: Hussain et al., 1983; Martinez and Sudre, 1995). This makes the complex of characters relating to ankle morphology useful for phylogenetic analysis of higher taxa. One of the problems with phylogenetic analyses based on tarsal morphology is the delineation of cladistic characters from a morphological continuum. Phylogenetic characters should be consistent and inde- pendent, and the delineation of morpho- logical characters greatly affects cladogram topology. We believe that functional studies greatly elucidate the way in which char- can acters are delineated and do so in this study. Central to our study is the shape of the artiodactyl astragalus. The astragalar mor- phology of artiodactyls is unique and plays an important role in locomotor function (Schaeffer, 1947, 1948). However, its unusual function is the result of several morpholog- ical features, a variable subset of which oc- casionally occurs in other mammals as well. Therefore, astragalar morphology cannot be considered a single character, a point recog- nized by Schaeffer (1948) but not by most subsequent authors. 21 Because of its importance in character- izing artiodactyls, astragalar morphology plays a crucial role in determining the phy- logenetic affinities of cetaceans. Cetaceans have been placed within artiodactyls on the basis of a large amount of recent molecular data (Gatesy et al., 1996; Gatesy, 1997, 1998; Hasegawa and Adachi, 1996; Shimamura et al., 1997; Milinkovitch et al., 1998). This view is commonly disputed by morpholog- ical studies (Prothero et al., 1988; Thewis- sen, 1994; Geisler and Luo, 1998; Luckett and Hong, 1998; O'Leary, 1998; see also Milinkovitch and Thewissen, 1997). Here, we describe fragmentary astragali for pa- kicetid and ambulocetid cetaceans (for dis- cussion of these families, see Thewissen et al., 1996, and Thewissen and Hussain, 1998). These bones were previously unknown in [PAGE BREAK] 22 SYSTEMATIC BIOLOGY primitive cetaceans and have only been mentioned in a short report (Thewissen et al., 1998). These bones fill a major gap in character matrices used to study the higher phylogeny of ungulates, making it possible to score cetaceans for several previously un- known characters in the tarsus. We here present a character analysis of the complex of joints between the mammalian astragalus, calcaneum, cuboid, and navic- ular (Fig. 1). The midtarsal joint (anatomi- cal terms are explained in Appendix 1) is of specific interest. This complex joint involves four bones; the trochleated head of the ar- tiodactyl astragalus is part of the midtarsal joint. Mobility at the midtarsal joint, how- ever, is determined not only by the shape of the joint facets that make up this joint but also their relation to the ectal, sustentacu- lar, and distal calcaneocuboid joints. These facets sometimes lock the bones that partic- ipate in the midtarsal joint at the point of contact between calcaneum and astragalus. Our description focuses on a primitive ungulate (Pleuraspidotherium), artiodactyls, and early whales, but we also consider the morphology of a broader range of placen- tals. There is relatively good consensus on the morphology of the primitive placental tarsus. Although different taxa were used to identify the placental morphotype by dif- ferent authors (Choeroclaenus by Schaeffer, 1947; Protungulatum by Szalay and Decker, 1974; Arctocyon by Cifelli, 1983), all are rel- atively similar in the characters as delin- eated here. The great age of these taxa and their retention of primitive traits are also consistent with their basal position on most cladograms. We use the astragalus of the ar- chaic ungulate Pleuraspidotherium (Thewis- sen, 1991) as a basis for our description and assume that most of its character scores are plesiomorphic with respect to modern un- gulates. This assumption is not critical for our results, however, because we do not con- struct a cladogram based on our limited data set; we only map our characters on existing cladograms. We scored characters for the fossil taxa listed in Table 1 as well as for several ex- tant forms. Scores are reported in Table 2, and the taxa used represent a cross-section VOL. 48 1999 THEWISSEN ET AI of all mammals as a reference for delineat- ing characters and to facilitate comparisons. We map these characters on a cladogram that has been proposed for ungulates, recog- nizing that, although these characters con- tribute to the phylogenetic resolution within ungulates, a complete phylogenetic analy- sis should rely on all available data, not just those of the ankle. Finally, we assess the tarsal data in light of the conflict between morphological and molecular studies of the ungulate phylogenetic tree. DESCRIPTION Midtarsal Joint The astragalar head of Pleuraspidotherium (Fig. 1b) is strongly convex mediolater- ally and dorsoplantarly. This permits some movement between astragalus and navicu- lar in all directions with no obvious axis of mobility. Movement at this joint is limited in part by the remainder of the midtarsal joint. The plane of the joint between calcaneum and cuboid in Pleuraspidotherium and other primitive mammals is more proximal than that between astragalus and navicular (Sza- lay and Decker, 1974), which is not the case in other placentals (Fig. 1a). The cuboid facet of the calcaneum is rounded in outline prim- itively, is mediolaterally and dorsoventrally concave, and faces distally. This mismatch in shapes between the astragalonavicular ar- ticulation on the one hand and the calca- neocuboid articulation on the other (Fig. 1b) limits the degree of mobility of the midtarsal joint. The cuboid does not articulate with the astragalus. The head of the astragalus in artiodactyls (Fig. 1d) has the shape of a broad trochlea (Schaeffer, 1947). This shape increases dor- soplantar mobility of the navicular on the astragalus. In addition, mediolateral mobil- ity is completely checked by a parasagittal groove that locks the navicular onto the as- tragalus. The calcaneocuboid joint of artiodactyls is also modified (Fig. 1d). It is narrow, re- stricted laterally, and slopes strongly dor- sodistally. This shape allows the cuboid to slide on the calcaneum while the navicu- lar moves on the astragalus and increases ASTRAGALUS CA CUNET FORMES NA (a) (f) (i) FIGURE 1. NV V -CU ECTAI SUST. (c SL DIST.AS Left tarsal bon astragalus and calcaneum of, tiodactyla), and Hyracotheriu (Mesonychia), Hyracotherium m) Dorsal view of astragalus Table 1. Drawings are not to numbers. Calc-cub= calcaneo facet. [PAGE BREAK] 1999 THEWISSEN ET AL. CETACEAN ANKLE MORPHOLOGY AND MAMMAL PHYLOGENY 23 ASTRAGALUS CALCANEUM CUNET FORMES (a) II III N V (b) NAVICULAR CUBOID TROCHLEA HEAD HEAD (c) ECTAL (d) CALC.-CUB.- (e) SUST. et 1- (f) (g) SUST. n r- a- b) .al (k) (h) he ls ea or- he ›il- tal as- yls re- lor- 1 to (CU- Ises (i) FIGURE 1. DIST.ASTR. (j) (1) HEAD (m) Left tarsal bones of placental mammals. (a) Tarsus of a dog in dorsal view. (b-e) Distal view of astragalus and calcaneum of, respectively, Pleuraspidotherium (condylarth), Oryctolagus (Lagomorpha), Capra (Ar- tiodactyla), and Hyracotherium (Perissodactyla). (f-j) Plantar view of astragalus of Pleuraspidotherium, Dissacus (Mesonychia), Hyracotherium, Diacodexis (Artiodactyla), and pakicetid cetacean (H-GSP 97227), respectively. (k- m) Dorsal view of astragalus of Dissacus, pakicetid, and Diacodexis, respectively. Specimen numbers are listed in Table 1. Drawings are not to scale and some are reversed from right elements. Roman numerals refer to digit numbers. Calc-cub= calcaneocuboid facet; dist astr= distal astragalar facet; ectal=ectal facet; sust= sustentacular facet. [PAGE BREAK] 24 SYSTEMATIC BIOLOGY TABLE 1. Fossil species used in this analysis, with museum number and literature reference. Order "Condylarth" Arctocyonia Phenacodonta Perissodactyla Artiodactyla Mesonychia Proboscidea Desmostylia Litopterna Cetacea Species Hyopsodus walcottianus Pleuraspidotherium aumonieri Arctocyon primaevus Tetraclaenodon puercensis Meniscotherium chamense Hyracotherium sp. Diacodexis pakistanensis Dissacus europaeus Pachyaena ossifraga Anthracobune pinfoldi Paleoparadoxia tubatai Asmithwoodwardia Pakicetid Ambulocetid Specimen number AMNH 14654 MNHN BR 10726, BR 55L MNHN CR 750, CR 752 AMNH 2468, AMNH 2493 USNM 18282, USNM 22918 AMNH 48663 H-GSP 5021, H-GSP 5274, GSP-UM 282 MNHN BR 21L USGS 25292 GSP-UM 1745 NMST-P-5601-45, UHR 18466-31 AMNH 109555 H-GSP 97227 H-GSP 96098, 97113 VOL. 48 Reference Gazin, 1968 Thewissen, 1991 Russell, 1964 Radinsky, 1966 Gazin, 1965 Thewissen and Hussain, 1990 Thewissen, 1991 O'Leary and Rose, 1995 Gingerich et al., 1990 Shikama, 1966 Cifelli, 1983 AMNH, American Museum of Natural History, New York; GSP-UM, Geological Survey of Pakistan-University of Michigan, Islamabad; H-GSP, Howard University, Geological Survey of Pakistan, Islamabad; NMST, National Science Museum, Tokyo; UHR, Hokkaido University collections, Hokkaido, Japan; MNHN, Museum National d'Histoire Naturelle, Paris, USGS, U. S. Geological Survey, Washington, D.C., USNM, U.S. National Museum, Smithsonian Institution, Washington, D.C. TABLE 2. Character matrix for tarsal characters described in text. oooooo Characters Taxon Hyopsodus Pleuraspidotherium Arctocyon Tetraclaenodon Meniscotherium Hyracotherium Diacodexis Sus Capra Hippopotamus Camelus Dissacus Pachyaena Anthracobune Paleoparadoxia Dendrohyrax Asmithwoodwardia Eocene cetaceans Canis Mustela Macaca Rattus Marmota Talpa Tupaia Oryctolagus Dasypus Didelphis boooo0000 000100010 12222323122121212. 100111001001¯¯¯o ooooooooo 1999 THEWISSEN ET AL.- dorsoplantar mobility of t The shape change in jo tragalus and navicular c between calcaneum and other occurs in all artio two changes do not alwa In lagomorphs (Oryctola litopterns (Asmithwoodwa Ernestokokenia: Cifelli, 198 distal astragalus has an e dorsoplantar mobility ba what trochleated facet sha neum and cuboid cannot in artiodactyls, thus limi excursion at the midtars logically, the calcaneocub sodactyls (Fig. 1e) is si artiodactyls in having la mobility, but the arrangem in the perissodactyl ankle the dorsoplantar plane. The astragalar head (Dissacus, Fig. 1g) is also t it is mediolaterally slightly soplantarly strongly conv that separates the parts of t ulate with cuboid and nav strongly oblique, as in per ing dorsoplantar mobility ilar crest is present in no dactyls, it extends dorsop not disrupt mobility in th icular and cuboid of run (Hussain et al., 1983) and The astragalar head of 97227, Fig. 1j and 1) is m slightly convex. The facet flattened. This shape all dorsoplantar and medio! less than in the rounded : a primitive mammal and the trochleated head of an Sustentacular The sustentacular fa dotherium (Fig. 1f) consi The larger part is oval proximal to the astragal. mediolaterally and sligh modistally. Continuous w proximal to it is a stron that is expanded onto th [PAGE BREAK] .5 ty al al n, 1910 1999 THEWISSEN ET AL. CETACEAN ANKLE MORPHOLOGY AND MAMMAL PHYLOGENY 25 dorsoplantar mobility of the midtarsal joint. The shape change in joints between as- tragalus and navicular on one hand and between calcaneum and cuboid on the other occurs in all artiodactyls but these two changes do not always occur together. In lagomorphs (Oryctolagus, Fig. 1c) and litopterns (Asmithwoodwardia, synonym of Ernestokokenia: Cifelli, 1983), the head of the distal astragalus has an expanded range of dorsoplantar mobility based on its some- what trochleated facet shape, but the calca- neum and cuboid cannot slide as they do in artiodactyls, thus limiting dorsoplantar excursion at the midtarsal joint. Morpho- logically, the calcaneocuboid joint of peris- sodactyls (Fig. 1e) is similar to that of artiodactyls in having laterally restricted mobility, but the arrangement of other facets in the perissodactyl ankle limits mobility in the dorsoplantar plane. The astragalar head of mesonychians (Dissacus, Fig. 1g) is also trochleated in that it is mediolaterally slightly concave and dor- soplantarly strongly convex. It bears a crest that separates the parts of the head that artic- ulate with cuboid and navicular. This crest is strongly oblique, as in perissodactyls, limit- ing dorsoplantar mobility. Although a sim- ilar crest is present in nonruminant artio- dactyls, it extends dorsoplantarly and does not disrupt mobility in this plane. The nav- icular and cuboid of ruminants are fused (Hussain et al., 1983) and no crest is present. The astragalar head of pakicetids (H-GSP 97227, Fig. 1j and 1) is mediolaterally only slightly convex. The facet is dorsoplantarly flattened. This shape allows very limited dorsoplantar and mediolateral mobility- less than in the rounded astragalar head of a primitive mammal and much less than in the trochleated head of an artiodactyl. Sustentacular Facet The sustentacular facet of Pleuraspi- dotherium (Fig. 1f) consists of two parts. The larger part is oval and located just proximal to the astragalar head; it is flat mediolaterally and slightly convex proxi- modistally. Continuous with this part and proximal to it is a strongly concave facet that is expanded onto the medial half of the plantar surface of the trochlea. The con- cavity of this part is oblique to the long axis of the bone and has the same axis as the concavity of the ectal facet. Combined, these two facets limit the mobility of the calcaneum on the astragalus to a direction oblique to the long axis of the foot. The sustentacular facet of Pleuraspidotherium is not continuous with the articular surface of the astragalar head. The sustentacular facet of plantigrade mammals, such as Pleuraspi- dotherium, has a weight-bearing function, transmitting force from the astragalus to the calcaneum. The sustentacular facet of artiodactyls (Fig. 1i) is greatly expanded. Its mediolateral profile may be concave or convex, but it is always convex proximodistally and the main axis of mobility is dorsoplantar. The sustentacular facet of artiodactyls covers most of the width of the plantar side of the astragalus. In mesonychians, the sustentacular facet is similar to that of Pleuraspidotherium, in that it consists of a concave and a convex part that limit mobility between astragalus and calcaneum. The sustentacular facet of pakicetid cetaceans (H-GSP 97227) is long and narrow, limited to the medial third of the astragalus, but it is proximodistally convex, as in artiodactyls. Only a small fragment of the sustentacular facet of the ambulo- cetid astragalus (H-GSP 96098) is preserved. Its morphology is consistent with that of the pakicetid. Ectal Facet The ectal facet of Pleuraspidotherium (Fig. 1f) faces plantarly and is elongate and strongly concave along an oblique axis. This concavity matches that of the sus- tentacular facet and locks the ectal facet of the astragalus onto the calcaneum, lim- iting mobility at the midtarsal joint. In perissodactyls (Fig. 1h), litopterns, and lagomorphs, the axis of the ectal facet ex- tends more mediolaterally than in prim- itive astragali (Fig. 1f), limiting dorso- plantar mobility between astragalus and calcaneum. The ectal facet of artiodactyls is highly modified. It is narrow and lies on the lateral [PAGE BREAK] 26 SYSTEMATIC BIOLOGY side of the calcaneum (and is hence not vis- is strongly reduced and variable in outline. ible in Fig. li). The ectal facet of artiodactyls This reduction permits astragalar rotation independent of the calcaneum (Schaeffer, 1947). The ectal facet of mesonychians (Fig. 1g) is similar to that of Pleuraspidotherium. The pakicetid astragalus does not preserve the ectal facet, but the proximal part of the am- bulocetid astragalus (H-GSP 96098) is well- preserved. Its ectal facet is laterally placed (see Thewissen et al., 1998). No part of the and resembles artiodactyls in morphology ectal facet extends on the plantar surface of the astragalus. FUNCTIONAL INFERENCE The artiodactyl midtarsal joint is uniquely mobile dorsoplantarly but is immobile mediolaterally as a result of extensive re- modeling of the ankle. This remodeling ap- pears to consist of four independent cladis- tic characters, none of which is unique to artiodactyls. First, the astragalar head is expanded dorsoplantarly (character 2, Ap- pendix 2), as is also the case in litopterns and mesonychians. Second, the plane of rotation of the astragalonavicular joint is dorsoplantar (character 3), as occurs also in lagomorphs and dasypodids. Third, the calcaneocuboid joint is narrow and ex- tends from proximoplantar to distodorsal on the lateral side of the foot (charac- ter 5), similar to perissodactyls. Fourth, the ectal facet is small and faces later- ally (character 8), as it does in primitive cetaceans. Artiodactyls share all four char- acters, but litopterns, mesonychians, lago- morphs, dasypodids, perissodactyls, and cetaceans have only some of the characters and their midtarsal joints are not highly mo- bile. Increased dorsoplantar mobility in artio- dactyls is combined with enhanced stability mediolaterally. This is related to two fea- tures of the heel. First, movement at the astragalar head is restricted mediolater- ally (character 1), as it is in perissodactyls, mesonychians, desmostylians, hyracoids, lagomorphs, and primitive cetaceans. Sec- ond, the sustentacular facet is expanded to VOL. 48 1999 THEWISSEN ET AL.-CET cover much of the plantar face of the astra- galus (character 6). This character is unique to artiodactyls. PHYLOGENETIC INFERENCE The modern orders of ungulates have a long independent history, and rever- sals and autapomorphies that accumulated since their divergence may seriously af- fect character analyses. The absence of the tarsus in modern cetaceans is an excel- lent example of such an autapomorphy. To avoid problems of homoplasy during their independent evolution, we studied as many Paleogene representatives of or- ders as possible. Because our set of char- acters is limited to those in the tarsus (Ta- ble 2), we did not execute a phylogenetic analysis, but instead mapped our charac- ters on a published phylogeny. We chose the phylogeny of Prothero et al. (1988) be- cause it includes nearly all of the rele- vant taxa and is fully resolved. Overall, this analysis is relatively similar to more broad-scale morphological analyses based on modern taxa (e.g., Gaudin et al., 1996). The Prothero et al. phylogeny (Fig. 2a) was not based on an explicit phyloge- netic analysis and thus may not be the most-parsimonious topology, but it is use- ful as a starting point for our study. Map- ping our tarsal data on this phylogeny indicates that 29 character state changes took place. Most notable among the char- acter changes implied in this phylogeny is character 8. The derived state of this character is the only feature that could be interpreted as a unique synapomor- phy of cetaceans and artiodactyls; instead, it is treated as a homoplasious in this analysis. Recent molecular data sets (reviewed by Gatesy, 1998) are incongruent with the topol- ogy of Figure 2a and suggest that cetaceans are allied with artiodactyls. Amending Fig- ure 2a to make Cete (Mesonychia plus Cetacea) the sister group of artiodactyls does not decrease the step length, but moving only Cetacea to the artiodactyl branch is more parsimonious (27 steps). Considering Mesonychia as the sister group to combined Cetacea plus Artiodactyla also costs 27 steps (a) 29 steps (c) 31 steps FIGURE 2. Four cladograms on w proposed by Prothero et al. (1988). St not affect character evolution of tars and mesonychians are here interprete and morphological studies. (c) Modit Choosing different sister groups for C with perissodactyls, cetaceans, meso is more-parsimonious than the othe calculated (by using MacClade, versi [PAGE BREAK] 1999 THEWISSEN ET AL-CETACEAN ANKLE MORPHOLOGY AND MAMMAL PHYLOGENY S VESTSS 15 g IS OS Dlacodexis Hippopotamus Sus Capra Camelus Arctocyon Hyopsodus cetacean Diacodexis Hippopotamus Sus Capra Camelus cotacoan Disaccus Pachysona Arctocyon 27 (a) 29 steps Dissacus Pachyaena Tetraclaenodon Plauraspidotherium Meniscotherium Anthracobune Paleoparadoxia Hyracotherium Dendrohyrax (b) 27 steps Hyopsodus Tetraclaenodon Pleuraspidotherium Meniscotherium Anthracobuna Paleoparadoxia Hyracotherium Dendrohyrax (c) 31 steps Diacodexis Hippopotamus Sus Capra Camelus cetacean Disaccus Pachyaena Arclocyon Hyopsodus Tetraclaenodon Pleuraspidotherium Meniscotherium Anthracobune Paleoparadoxia Hyracotherium Dandrohyrax Diacodexis Hippopotamus Sus Capra Camelus cetacean Disaccus Pachyaena Hyracotherium Arctocyon Hyopsodus Tetraclaenodon Plouraspidothorium Moniscolherium Anthracobune (d) 25-26 steps Paleoparadoxla Dendrohyrax FIGURE 2. Four cladograms on which the tarsal data were mapped. (a) Cladogram of ungulate relations as proposed by Prothero et al. (1988). Step length is unaffected by fully resolved artiodactyl relations, and this does not affect character evolution of tarsal characters. (b) Somewhat modified Prothero et al. cladogram. Cetaceans and mesonychians are here interpreted as successive sister groups of artiodactyls, consistent with some molecular and morphological studies. (c) Modified Prothero et al. cladogram, in which Cete is included within Artiodactyla. Choosing different sister groups for Cete does not affect step length. (d) Additional modifications of the cladogram with perissodactyls, cetaceans, mesonychians, and artiodactyls as an unresolved polychotomy. This cladogram is more-parsimonious than the other cladograms and is consistent with some molecular data; step length was calculated (by using MacClade, version 3) on the basis of fully resolved equally parsimonious cladograms. [PAGE BREAK] 28 SYSTEMATIC BIOLOGY derived character linking artiodactyls to (Fig. 2b). Character 8 is the only uniquely cetaceans (to the exclusion of mesonychi- character 5 in Fig. 2b, where it is a homo- ans), but this grouping is also supported by plasy in perissodactyls. The same two char- acters also support the inclusion of Cetacea in Artiodactyla at a branch above Diacodexis, (e.g., Gatesy, 1997; Hasegawa and Adachi, (Fig. 2c) as suggested by molecular studies 1996; Shimamura et al., 1997). In this sce- nario, the astragalar head of cetaceans is in- terpreted as secondarily flattened (character 2). Thus, tarsal evidence does not support the monophyly of Cete (Fig. 2c). Molecular data (e.g., Lavergne et al., 1996) also suggest that Perissodactyla are not re- lated to tethytheres and hyracoids, contrary to what morphological studies suggest (e.g., Fischer, 1986; Novacek, 1992; Thewissen and Domning, 1992). Stanhope et al. (1996) supported close phylogenetic ties between Paraxonia (dolphin, pig, cow) and Perisso- dactyla. Making this modification (Fig. 2d) reduces the step length to 25 or 26. Our tarsal data are thus consistent with results of the molecular analysis of Stanhope et al. (1996). CONCLUSIONS Our character analysis suggests that the is a character complex consisting of four "trochleated astragalar head" of artiodactyls independent cladistic characters. Together these characters work to increase dorsoplan- tar mobility of the tarsus. Limiting mediolat- eral mobility is another functional change in the artiodactyl astragalus, and this is charac- terized by two cladistic characters. No mam- mals except artiodactyls combine the six (or even the initial four) characters, but only one of the six characters (expanded sustentacu- lar facet) is unique to artiodactyls. This facet is narrow in cetaceans but wide in primi- tive placentals. In artiodactyls, on the other hand, the sustentacular facet is expanded to cover the entire width of the plantar sur- face. Cetaceans and artiodactyls share the derived position and shape of the ectal facet, a condition not shared by mesonychians. We conclude that tarsal data support close phylogenetic ties between Cetacea, Artio- VOL. 48 dactyla, Mesonychia, and Perissodactyla, to the exclusion of the paenungulates. This is consistent with recent molecular data. New evidence of Eocene cetacean tarsal mor- phology is also consistent with inclusion of cetaceans in artiodactyls, if one assumes that the wide arc of rotation of the trochleated head was lost during the origin of Cetacea. Tarsal data form only a small part of the to- tal body of evidence bearing on cetacean re- lations, but it has been considered critical in evaluating artiodactyl-cetacean relations (Luckett and Hong, 1998). The tarsal data do not support mesonychian-cetacean rela- tions. ACKNOWLEDGMENTS We thank Tony Friscia and Lois Roe, the collectors of the two cetacean astragali described here. Richard Cifelli provided us with a collection of casts of tarsal bones, Ken Rose gave access to the Pachyaena tarsals, and Philip Gingerich gave access to the Anthracobune astragalus. Other specimens studied are at the Geo- logical Survey of Pakistan and the Museum National d'Histoire Naturelle in Paris. Fieldwork and analysis were supported by an NSF grant to J. G. M. T. REFERENCES CIFELLI, R. L. 1983. Eutherian tarsals from the late Paleocene of Brazil. Am. Mus. Novit. 2761:1-31. FISCHER, M. S. 1986 Die Stellung der Schliefer (Hyra- coidea) im phylogenetischen System der Eutheria. Cour. Forschungsinst. Senckenb. 84:1-132. GATESY, J. 1997. More DNA support for a Cetacean/ Hippopotamidae clade: The blood-clotting protein gene y-fibrinogen. Mol. Biol. Evol. 14:537-543. GATESY, J. 1998. Molecular evidence for the phyloge netic affinities of Cetacea Pages 63-111 in The emer- gence of whales: Evolutionary patterns in the origin of Cetacea (J. G. M. Thewissen, ed.). Plenum, New York. GATESY, J., C. HAYASHI, M. A. CRONIN, AND P. ARCTANDER. 1996. Evidence from milk casein genes that cetaceans are close relatives of hippopotamid ar- tiodactyls. Mol. Biol. Evol. 13:954-963. GAUDIN, T. J., J. R. WIBLE, J. A. HOPSON, AND W. D. TURNBULL. 1996. Reexamination of the morpholog- ical evidence for the cohort Epitheria (Mammalia, Eutheria). J. Mammal Evol. 3:31-77. GAZIN, C. L. 1965. A study of the early tertiary condy- larthran mammal Meniscotherium. Smithson. Misc. Collect. 149:1-98. GAZIN, C. L. 1968. A study of the Eocene condy- larthran mammal Hyopsodus. Smithson. Misc. Col- lect. 153:1-90. GEISLER, J. H., AND Z. Luo. 1998. Relationships of Cetacea to terrestrial ungulates and the evolution of cranial vasculature in Cete. Pages 163-212 in The 1999 THEWISSEN ET AL.- emergence of whales: Evolutio origin of Cetacea (J. G. M. Thev New York. GINGERICH, P. D., D. E. RUSSELL, 1990. Astragalus of Anthrac Proboscidea) from the early. Kashmir. Contrib. Mus. Pale 28:71-77. HASEGAWA, M., AND J. ADACHI. 19 sition of cetaceans relative to art: of mitochondrial and nuclear s Evol. 13:710-717. HUSSAIN, S. T., P. Y. SONDAAR, G. M. THEWISSEN, E. F. H. M. SPOOR. 1983. Fossil mammal field atlas. Part I: The artiodact Geol. Surv. Pak. 14:1-14. LAVERGNE, A., E. DOUZERY, T. STICH AND M. S. SPRINGER. 1996. malian relationships: Evidenc monophyly is provided by c drial 125 rRNA sequences. Mo 6:245-258. LUCKETT, W. P., AND N. HONG. 19° lationships between the order Cetacea: A combined assessm ical and molecular evidence. 5:127-182. MARTINEZ, J.-N., AND J. SUDRE. 1 of Paleogene artiodactyls: Co ogy, variability and prediction o 28.197-209. MATTHEW, W. D. 1937. Paleocen Juan Basin. Trans. Am. Philos. S MILINKOVITCH, M. C., M. BÉRUBÉ, 1998. Are cetaceans highly d Pages 113-131 in The eme Evolutionary patterns in the (J. G. M. Thewissen, ed.). Plenu: MILINKOVITCH, M. C., AND J. G. M Even-toed fingerprints on wha 388:622-624. NOVACEK, M. J. 1992. Fossils, top and the higher level phylogeny mals. Syst. Biol. 41:58-73. O'LEARY, M. A. 1998. Phylogenet: reassessment of the dental evic chian and cetacean clade. Pages gence of whales: Evolutionary of Cetacea (J. G. M. Thewissen York. O'LEARY, M. A., AND K. D. Ros skeleton of the early Eocene m (Mammalia, Mesonychia). J. V 430. PROTHERO, D., E. MANNING, AND M phylogeny of ungulates. Pages logeny and classification of the Mammals (M. Benton, ed.). Clar England. RADINSKY, L. B. 1966. The adapt phenacodontid condylarths an Perissodactyla. Evolution 20:40 [PAGE BREAK] KLINE SCIENCE LIB 1999 THEWISSEN ET AL. CETACEAN ANKLE MORPHOLOGY AND MAMMAL PHYLOGENY emergence of whales: Evolutionary patterns in the origin of Cetacea (J. G. M. Thewissen, ed.). Plenum, New York GINGERICH, P. D., D. E. RUSSELL, AND N. A. WELLS. 1990. Astragalus of Anthracobune (Mammalia, Proboscidea) from the early-middle Eocene of Kashmir. Contrib. Mus. Paleontol. Univ. Mich. 28 71-77. HASEGAWA, M., AND J. ADACHI. 1996. Phylogenetic po- sition of cetaceans relative to artiodactyls: Reanalysis of mitochondrial and nuclear sequences. Mol. Biol. Evol. 13:710-717. HUSSAIN, S. T., P. Y. SONDAAR, S. M. I. SHAH, J. G. M. THEWISSEN, E. F. H. M. COUSIN, AND C. F. SPOOR. 1983. Fossil mammal bones of Pakistan, a field atlas. Part I: The artiodactyl astragalus. Mem. Geol Surv. Pak. 14:1-14. LAVERGNE, A., E. DOUZERY, T. STICHLER, F. M. CATZEFLIS, AND M. S. SPRINGER. 1996. Interordinal mam- malian relationships: Evidence for paenungulate monophyly is provided by complete mitochon- drial 125 rRNA sequences. Mol. Phylogenet. Evol 6:245-258. LUCKETT, W. P., AND N. HONG. 1998. Phylogenetic re- lationships between the orders Artiodactyla and Cetacea: A combined assessment of morpholog- ical and molecular evidence. J. Mammal Evol. 5:127-182. MARTINEZ, J.-N., AND J. SUDRE. 1995. The astragalus of Paleogene artiodactyls: Comparative morphol- ogy, variability and prediction of body mass Lethaia 28:197-209. MATTHEW, W. D. 1937. Paleocene faunas of the San Juan Basin. Trans. Am. Philos. Soc., N. S. 30:1-510. MILINKOVITCH, M. C., M. BÉRUBÉ, AND P. J. PALSBØLL. 1998. Are cetaceans highly derived artiodactyls? Pages 113-131 in The emergence of whales: Evolutionary patterns in the origin of Cetacea (J.G M. Thewissen, ed.). Plenum, New York. MILINKOVITCH, M. C., AND J. G. M. THEWISSEN. 1997. Even-toed fingerprints on whale ancestry. Nature 388.622-624. NOVACEK, M. J. 1992. Fossils, topologies, missing data, and the higher level phylogeny of eutherian mam- mals. Syst. Biol. 41:58-73. O'LEARY, M. A. 1998. Phylogenetic and morphometric reassessment of the dental evidence for a mesony- chian and cetacean clade Pages 133-162 in The emer- gence of whales: Evolutionary patterns in the origin of Cetacea (J. G. M. Thewissen, ed.). Plenum, New York. O'LEARY, M. A., AND K. D. ROSE. 1995. Postcranial skeleton of the early Eocene mesonychid Pachyaena (Mammalia, Mesonychia). J. Vert. Paleont. 15:401- 430. PROTHERO, D., E. MANNING, AND M. FISCHER. 1988. The phylogeny of ungulates. Pages 201-234 in The phy- logeny and classification of the tetrapods, Volume 2. Mammals (M. Benton, ed.). Clarendon Press, Oxford, England. RADINSKY, L. B. 1966. The adaptive radiation of the phenacodontid condylarths and the origin of the Perissodactyla. Evolution 20:408–417. 29 RUSSELL, D. E. 1964. Les mammiféres paléocènes d'Europe. Mém. Mus. Natl. d'Hist. Nat. Ser. C 13:1- 324. SCHAEFFER, B. 1947. Notes on the origin and func- tion of the artiodactyl tarsus. Am. Mus. Novit. 1350:1-24. SCHAEFFER, B. 1948. The origin of a mammalian ordi- nal character. Evolution 2:164-175. SHIKAMA, T. 1966. Postcranial skeletons of Japanese Desmostylia, Palaeontol. Soc. Jpn. Spec. Pap. 12:1- 202. SHIMAMURA, M, H. YASUE, K. OHSHIMA, H. ABE, H. KATO, T. KISHRO, M. GOTO, I. MUNECHIKAL, AND N. OKADA. 1997. Molecular evidence from retro- posons that whales form a clade within even-toed ungulates. Nature 388:666-670. STANHOPE, M. J., M. R. SMITH, P. G. WADDELL, C. A. PORTER, M. S. SHIVJI, AND M. GOODMAN. 1996. Mam- malian evolution and the interphotoreceptor retinoid binding protein (IRBP) gene: Convincing evidence for several superordinal clades. J. Mol. Evol. 43:83-92. SZALAY, F. S., AND R. L. DECKER. 1974. Origins, evo- lution of the tarsus in late Cretaceous eutheria and Paleocene primates. Pages 223-259 in Primate Lo- comotion (F. A. Jenkins, ed.). Academic Press, New York. SZALAY, F S., AND G. DRAWHORN. 1980. Evolution and diversification of the Archonta in an arboreal milieu. Pages 133-169 in Comparative biology and evolu- tionary relationships of tree shrews (W. P. Luckett, ed.). Plenum, New York. TASSY, P. 1996. Who is who among Proboscidea? Pages 39-54 in The Proboscidea: Evolution and palaeoecology of elephants and their relatives (J. Shoshani and P. Tassy, eds.). Oxford Univ. Press, New York. THEWISSEN, J. G. M. 1991. Limb osteology and func- tion of the primitive Paleocene ungulate Pleuraspi- dotherium with notes on Tricuspiodon and Dissacus (Mammalia). Geobios 24:483–495. THEWISSEN, J. G. M. 1994 (published in 1995). Phylo- genetic aspects of cetacean origins: A morphological perspective. J. Mammal. Evol. 2.157-184. THEWISSEN, J. G. M., AND D. P. DOMNING. 1992. The role of phenacodontids in the origin of the modern orders of ungulate mammals. J. Vertebr. Paleontol. 12:494- 504. THEWISSEN, J. G. M., AND S. T. HUSSAIN. 1990. Postcra- nial osteology of the most primitive artiodactyl, Dia- codexis pakistanensis (Dichobunidae). Anat. Hist. Em- bryol. (Berl.) 19.37-48. THEWISSEN, J. G. M., AND S. T. HUSSAIN. 1998. Sys- tematic review of the Pakicetidae, early and mid- dle Eocene Cetacea (Mammalia) from Pakistan and India. Pages 220-238 in Dawn of the age of mammals in Asia (C. Beard and M. R. Dawson, eds.). Carnegie Museum of Natural History, Pittsburgh, Pennsylvania. THEWISSEN, J. G. M., S. I. MADAR, AND S. T. HUSSAIN. 1996. Ambulocetus natans, an Eocene cetacean (Mammalia) from Pakistan. Cour. Forschungsinst. Senckenb. 191:1-86. of n e [PAGE BREAK] 30 SYSTEMATIC BIOLOGY THEWISSEN, J. G. M., S. I. MADAR, AND S. T. HUSSAIN. Nature 395:452. 1998. Whale ankles and evolutionary relationships. Received 21 July 1998; accepted 15 September 1998 Associate Editor: P. Waddell APPENDIX 1 EXPLANATION OF SOME ANATOMICAL TERMS site to the sole. Dorsal side of foot. The side of the foot that is oppo- Ectal facet (posterior calcaneoastragalar facet). The facet on the astragalus that articulates with the prox- imolateral side of the calcaneum. It is positioned on the plantar surface of the astragalus in most mammals (Fig. 1f), but it is lateral in artiodactyls. Facet (or joint facet). A surface on a bone that artic- ulates with another bone. Two contacting facets of two bones make up a joint. Head of astragalus. Distal joint of astragalus (with navicular and sometimes also cuboid), which usually (primitively) has a rounded, convex shape (the ball of a ball-and-socket joint, convex in all directions, Fig. 1b and c). The astragalar head of artiodactyls does not have this shape but instead bears a hinge joint. The latter is called the trochleated astragalar head (Fig. 1d). Midtarsal joint. The joint between the calcaneum and astragalus on one side and the cuboid and navicular on the other. Plantar side. The side of the foot where the sole is. Sustentacular facet. The articular surface on the plan- tar part of the astragalus (Fig. 1f and g) that contacts the medially projecting part of the calcaneum (sustentacu- lum tali). Trochlea. Hinge joint, allowing great mobility in one direction, and very little in the direction perpendicular to that. In the case of the astragalus, the trochleated as- tragalar head enlarges dorsoplantar mobility and limits mediolateral mobility (Fig. 1b and c). APPENDIX 2 CLADISTIC CHARACTERS OF TARSUS The relations among astragalus, calcaneum, navic- ular, and cuboid can be described as cladistic charac- ters. We consider these characters independent because combinations of them occur in a variety of other mam- mals, and no two characters are functionally depen- dent in an obvious way. Character state (0) is considered primitive for placentals. VOL. 48 1. Head of the astragalus is mediolaterally strongly convex (0) or flat/concave (1). The derived state restricts adduction and abduction at the mid- tarsal joint. The shape of the astragalar facet galus. of the navicular will match that of the astra- 2. Head of the astragalus describes a small-angled arc dorsoplantarly (0) or extends through a wide arc (1). The derived state indicates increased dor- soplantar mobility of the navicular on the as- tragalus. This character can be quantified. In ar- tiodactyls, the navicócuboid rotates through an arc of ~200, similar to mesonychians (Disaccus: 200°) and litopterns (Asmithwoodwardia: 210). Prim- itively the condyle rotates through a smaller an- gle (Pleuraspidotherium 185°, Anthracobune: 105°), whereas in cetaceans this angle is very small (pa- kicetid: 60°) 3. Plane in which the astragalonavicular joint rotates extends dorsoplantar with respect to the plane of the trochlea (1) or extends at an oblique axis to the head (2), or there is no rotational plane because the facet is nearly flat (3). Rotation at the astragalonavic- ular joint is restricted to one plane in a great number of taxa, but the plane of this motion differs. In taxa where the astragalar head is a condyle (the primi- tive state), this character cannot be scored. This is an unordered character. 4 Head of astragalus and cuboid do not (0) or do (1) articulate. 5. Joint between calcaneum and cuboid is oval and fac- ing distally (0) or elongate and oblique to the dorso- plantar direction of the calcaneum (1). The derived character state is necessary to allow sliding between these two bones during movement of the navicular on the astragalus. The size of the calcaneocuboid joint is inversely related to the size and position of the astragalar head (realigned longitudinal axis of the astragalar head in the terminology of Schaeffer, 1947). Thus, this character can also be scored if only an astragalus is available. 6. Sustentacular facet of astragalus is oval and posi- tioned on the medial half of the astragalus (0), or elongate and positioned on the medial third of the astragalus (1), or rectangular and covering nearly all of the astragalar width (2). This is an unordered character, mirrored by the sustentacular facet of the calcaneum. 7. Distal calcaneal facet of astragalus is indistinct from sustentacular facet (0) or clearly distinct (1). 8. Ectal facet is concave and faces plantarly (0) or faces laterally and is flat (1). 9. Articulation between calcaneum and fibula is ab- sent (0) or present (1). Syst. Biol 48(1).31-53, 1999 Using Novel Phyl Including Amin to Detect Inter. to the Posi. PETER J. WADDI 'Institu : Sci Abstract. We look at t mtDNA sequences of m lationships. To this end, better-established meth frequencies are changin LogDet amino acid dist with bootstrapping and models. To weight the n the data by using "site The bootstrap support! can claim unanimous su earlier branching patter root. The tRNA genes, fc versus all other sequenc data. A grouping of all amino acid data, and ro rejection of the older ta: Fereuungulata are defin timal tree for tRNA vers indicate the test is tendi placement tests suggest. support elephant and ar of Xenarthra and Africa subtree. Thus, while cast interesting new possibil mammal phylogeny; mi Generally, the molecu malian interordinal relati to be in closer agreem other than with the morph Springer et al., 1997). H some possible conflicts b drial (mt) DNA and nucl haps even within the mtl fying, then resolving, phy Is a major path for molec studies to advance along. An important questior whether a major data pa Present address: Institute of Massey University, Palmerston Email: waddell@onyx.si.edu




