Box 4
Folder 27. Archeotes History Zeuglodon
Item 3. Academic Articles






















Transcribed Text (OCR)
GARY MANGIACOPA ARCHIVE
============================================================
Title: B4F27I3
Slug: b4f27i3
Categories: Cryptozoology
Source: https://garymangiacopraarchive.com/b4f27i3
Pages: 24 scanned, 24 extracted
OCR: Google Vision API (document_text_detection)
Processed: 2026-06-06
============================================================
S.C.3.C
y
MONOPHYLY OR DIPHYLY IN THE ORIGIN OF WHALES
LEIGH VAN VALEN
Department of Anatomy, University of Chicago
Received January 20, 1967
The phyletic unity of the Cetacea is still
a matter of dispute. Recently Yablokov
(1964; see also Kleinenberg, 1958, 1959)
has given well-reasoned arguments that
much of the aquatic adaptation of the two
recent suborders was evolved separately,
from an ancestor without these adaptations.
Although not always compelling and some-
times overstated, these arguments are
persuasive and they may in general be ac-
cepted. For a review of the earlier litera-
ture on this subject see Kellogg (1936).
Yablokov and Kleinenberg further con-
clude, in agreement with several previous
authors, that the Odontoceti and Mysticeti
originated separately from terrestrial ani-
mals. Granting that much of the evolution
of whales is homoplastic, as is true for
probably all other mammalian orders ex-
cept the least diverse, it is still legitimate
to ask whether the latest common ancestor
of the recent suborders was sufficiently
specialized to be called a whale. The evi-
dence from recent forms is of little help
here, but the earliest fossil whales are more
promising. It is immediately obvious that
the better-known families of archaeocetes,
the Basilosauridae and Dorudontidae,
could not have given rise to any recent
whale, but it is not so obvious that the
Protocetidae could not have done so.
THE EARLIEST FOSSIL WHALES
Other than a partial scapula from the
London Clay (Tarlo, 1964), which simply
shows that whales existed in the early
Eocene but which was unfortunately given
a name (Anglocetus), the earliest and most
primitive known whales are the possibly
congeneric forms Protocetus and Pappo-
cetus from the middle Eocene of Africa
(with a referred vertebra from Texas).
Protocetus was well described and discussed
by Fraas (1904) and Kellogg (1936); it
EVOLUTION 22:37-41. March, 1968
37
is known from a skull without the mandi-
ble, and some vertebrae and ribs. Pappo-
cetus, for which see Andrews (1920), Kel-
logg (1936), and Van Valen (1966), is
known only from two mandibles and a
fragmentary axis. The somewhat later
genus Eocetus, also placed in the Proto-
cetidae, is clearly related to the Basilosaur-
idae and could equally well be referred to
the latter family.
Pappocetus came from the type locality
of the Ameki Formation in the Eastern
Region of Nigeria (Reyment, 1965). In
June, 1965, Reyment saw (but did not
collect because of inadequate equipment)
"many bones of a whale in situ" at the
type locality of the Ameki Formation, in
the middle part of its upper unit. This
find would be of about middle Lutetian
age (Reyment, letter of July 19, 1966),
which is presumably about the age for the
type specimen of Pappocetus also.
I have recently given reasons for believ-
ing that the Cetacea arose from a primitive
member of the Mesonychidae in about the
middle or late Paleocene (Van Valen, 1966:
90-93). Although some mesonychids were
secondarily carnivorous (Szalay and Gould,
1966:165-166), they belong phyletically to
the Ungulata (Van Valen, 1966:103).
REASONS FOR DIPHYLY
Most of the reasons given by Yablokov
(1964) for separate origin of the Odonto-
ceti and Mysticeti are such as cannot be
determined from the available fossils; in-
deed, most are from rarely preserved soft
structures and even biochemistry and be-
havior. Some, however, are osteological,
and these may be listed as follows.
1) Although the external nares of both
recent suborders are far back on the skull,
with concomitant retraction of the nasal
bones, the arrangements of the posterior
[PAGE BREAK]
JOURNAL OF PALEONTOLOGY, V. 49, No. 4, P. 745-751, 3 TEXT-FIGS., JULY 1975
Copyright 1975, The Society of Economic Paleontologists and Mineralogists
Geol. Soc. Amer. Bull. s
69. Equitability indices
pecies count. Ecology. 50 v
nd H. L. Sanders. 1969
of environmental predict
ty. Brookhaven Symposi
in, L. P. Atkin, and
ydrographic pre
North Carolina
Research. 18420
f Engineers. 1961 Outer
Inlet and Beaufort Inlet,
rmy Corps of Engineers.
Army Engineer District
Carolina, 26 p, appendices
use of Representatives
hore line, beach erosion
th, 2nd sess, House 1.
966. Numerical analysts
nges on the extra-tropical
helf. Limnol. Occ mgr 1
urce supply and
4:51-61.
: tectonics and illow
demism, an actualistic
-264.
1971. Climatic implication
stracode assemblage from
US. Geol. Surv. Prod
1 R. I. Wicklund. 1968 M
structure from the Florid
mer. Geogr. Soc., Serial
ient, Folio 15.
ED DECEMBER 6, 1974
IT RECEIVED FER: KY 18,
T'
A FOSSIL BEAKED WHALE (CETACEA: ZIPHIIDAE)
FROM THE MIOCENE OF KENYA
JAMES G. MEAD
National Museum of Natural History, Smithsonian Institution, Washington, D.C., 20560
ABSTRACT-A fossil beaked whale was discovered in 1964 in late middle Miocene sedi-
ments of northern Kenya. The specimen consists of a weathered rostral fragment, found in
association with a freshwater-terrestrial fauna. It is similar to the Miocene genera
Proroziphius and Belemnosiphius in lacking premaxillary basining. Those genera appear
to be related to such modern forms as Mesoplodon, whereas the remainder of the Miocene
ziphiids are probably more closely related to the living Ziphius Belemnoziphius com-
pressus Huxley, 1865, is designated the type species of the genus Belemnoziphius.
INTRODUCTION
ite first cetaceans, found in the lower
Eocene, are members of the suborder
coceti. Archaeocetes flourished in the
Focene, probably giving rise to the modern
orders of Cetacea during that time. The
xene presents a gap in the history of the
cea, yielding a few remnants of the ar-
cetes and some poorly known and aberrent
Some of these are intermediate be-
**** archaeocetes and the modern cetaceans.
the Miocene, however, cetacean remains are
lant, and members of most of the living
les and even a few living genera are pres-
Notable among these are the beaked
family Ziphiidae. This family is char-
ded by elongation of the rostrum, reduc-
of the dentition and elevation of the
region. A few of the early forms as-
ed to this family preserve characters link-
them to the more primitive squalodont
The specimen on which this report is based
of Comparative Zoology, Harvard
nity, Field No. 14-64K) was recovered
The author during the summer of 1964,
The un an expedition to the northern part of
under the leadership of Bryan Patter-
of Harvard University, supported by Na-
Science Foundation grant No. G. P.
The specimen was obtained from the
part of the Turkana Grit, about 20 feet
a basalt dated by K/Ar analysis at 16.7
13 million years (Bryan Patterson, personal
- 1969). This suggests a late middle
(Burdigalian) age. Maglio (1969)
used a more extended correlation of these
The locality is near Loperot, about
south-southeast of the town of Lod-
the Turkana District of Kenya (ap-
745
proximate geographic coordinates: 35° 50' E,
02° 20' N). The specimen was found in a
well indurated bed of coarse sand to fine con-
glomerate, associated with the following fauna:
bivalves, gastropods, Protopterus, Polypterus,
miscellaneous other fish, Tomistoma, unidenti-
fied crocodile remains, trionychid, testudinid
and other chelonian remains, hominoid, rodent,
creodont, mastodont, gomphothere, hyracoid,
suid, anthracothere, Dorcatherium and bovid
remains (Maglio, 1969; Bryan Patterson, per-
sonal commun., 1969). D. Van Damme (per-
sonal commun., 1973) has indicated that the
molluscan fauna associated with this cetacean
suggests "a swampy flood plain or swamps
connected with a periodically fluctuating river
system."
The cetacean specimen consists of the ros-
trum and the ascending processes of the max-
illae and premaxillae, broken from the rest of
the skull just anterior to the nares. The overall
length of the specimen is 82 cm, the greatest
width (posteriorly) is 55 cm. The specimen
contains the greater portions of the maxillae,
premaxillae and vomer.
The tip of the rostrum is slightly eroded on
the left side, but appears to be entire on the
right side (Text-fig. 1). The premaxillae are
separate at this point, forming a terminal notch
as seen in the living ziphiids. One of the prob-
lems with this particular specimen is determin-
ing the relative extent of the premaxillae and
vomer. On the dorsal surface the premaxillae
do not appear to have met in the midline.
About 5 cm posterior to the terminal notch,
the dorsal surface bears a median groove and
faint indications of paired grooves about 1.5
cm lateral to the median groove. The median
groove could be interpreted as the junction
of the premaxillae in the midline, or it could
[PAGE BREAK]
CENE CE
306
Pa.
Na.
-Fr.
Pmx.
Max.
Ant. n.
La.
S. or pr.
S. oc.
C.
Zyg.
Sq.
SYSTEMATIC ZOOLOGY
S. oc.
Pmx.
Max.
Fr
maxilla,
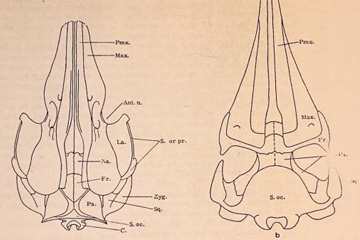
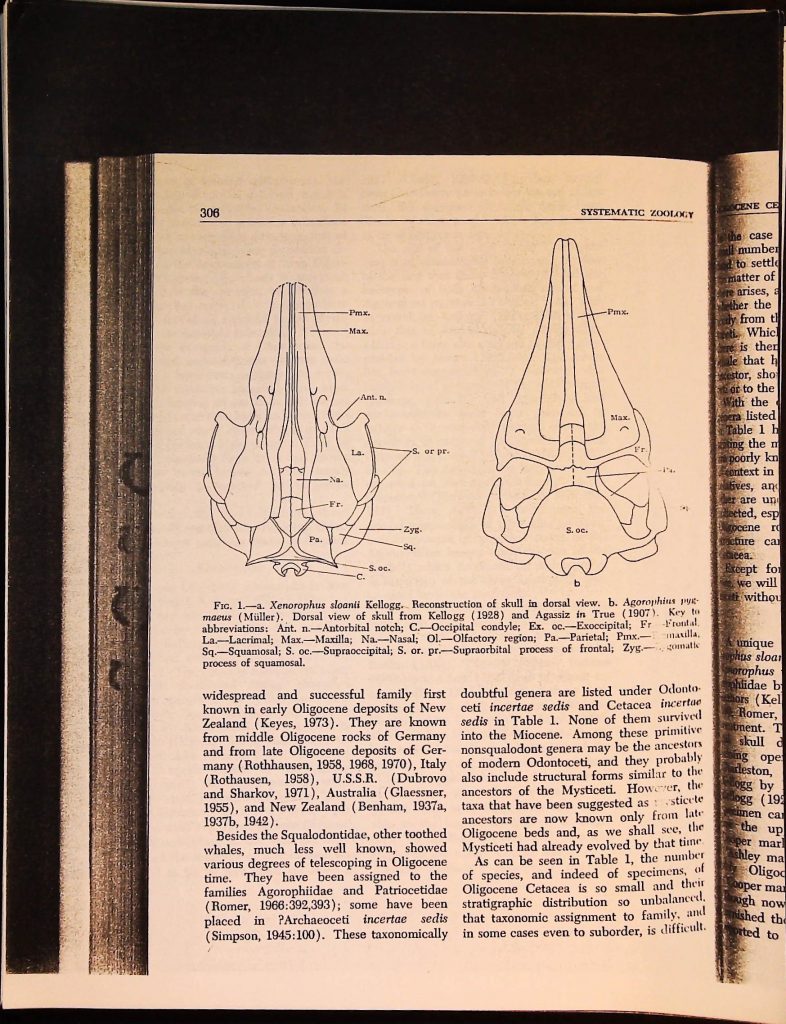
FIG. 1. a. Xenorophus sloanii Kellogg. Reconstruction of skull in dorsal view. b. Agorophius pyg-
maeus (Müller). Dorsal view of skull from Kellogg (1928) and Agassiz in True (1907). Key to
abbreviations: Ant. n.-Antorbital notch; C.-Occipital condyle; Ex. oc.-Exoccipital; Fr Frontal.
La. Lacrimal; Max.-Maxilla; Na.-Nasal; Ol.-Olfactory region; Pa.-Parietal; Pmx.-
Sq.-Squamosal; S. oc.-Supraoccipital; S. or. pr.-Supraorbital process of frontal; Zyg-gomatic
process of squamosal.
widespread and successful family first
known in early Oligocene deposits of New
Zealand (Keyes, 1973). They are known
from middle Oligocene rocks of Germany
and from late Oligocene deposits of Ger-
many (Rothhausen, 1958, 1968, 1970), Italy
(Rothausen, 1958), U.S.S.R. (Dubrovo
and Sharkov, 1971), Australia (Glaessner,
1955), and New Zealand (Benham, 1937a,
1937b, 1942).
Besides the Squalodontidae, other toothed
whales, much less well known, showed
various degrees of telescoping in Oligocene
time. They have been assigned to the
families Agorophiidae and Patriocetidae
(Romer, 1966:392,393); some have been
placed in PArchaeoceti incertae sedis
(Simpson, 1945:100). These taxonomically
doubtful genera are listed under Odonto-
ceti incertae sedis and Cetacea incertae
sedis in Table 1. None of them survived
into the Miocene. Among these primitive
nonsqualodont genera may be the ancestors
of modern Odontoceti, and they probably
also include structural forms similar to the
ancestors of the Mysticeti. However, the
taxa that have been suggested as
ancestors are now known only from late
Oligocene beds and, as we shall see, the
Mysticeti had already evolved by that time.
ysticete
As can be seen in Table 1, the number
of species, and indeed of specimens, of
Oligocene Cetacea is so small and their
stratigraphic distribution so unbalanced.
that taxonomic assignment to family, and
in some cases even to suborder, is difficult.
the case
number
to settle
matter of
re arises, a
ther the
ly from th
Whic
is ther
le that h
estor, sho
or to the
With the
era listed
Table 1 h
ing the m
poorly kn
ontext in
ves, and
er are un
Sceted, esp
Ocene I
racture ca
cea
Except for
we will
withou
unique
phus sloa
orophus
tidae b
ors (Kel
Romer,
ment. T
skull a
ng ope
arleston,
ogg by
Pgg (192
men ca
the up
per marl
shley ma
Oligo
oper ma
ugh now
ished the
orted to
[PAGE BREAK]
EMATIC ZOOLOGY
Pmx.
Max.
Fr.
Pa.
b. Agorophius P
le (1907). Key to
pital; Fr.-Frontal;
; Pmx.-Premaxilla,
d; Zyg.-Zygomatic
d under Conto-
Cetacea incertac
of them survived
g these primitive
y be the ancestors
nd they probably
ms similar to the
ti. However, the
ested as mysticete
n only from late
we shall see, the
lved by that time.
le 1, the number
of specimens, of
small and their
so unbalanced,
nt to family, and
border, is difficult.
COCENE CETACEA
the case of familial assignments, the
number of species involved makes it
to settle on definitive characters. In
matter of assigning genera to suborders,
arises, among other questions, that of
her the Mysticeti were descended di-
from the Archaeoceti or from Odon-
Whichever hypothesis is accepted,
is then the question of whether a
that has teeth, if it is a mysticete
stor, should be assigned to the Mysti-
to the ancestral suborder.
th the exception of Chonecetus, the
listed under Cetacea incertae sedis
Table 1 have been advanced as repre-
ng the mysticete ancestral type. Most
poorly known; all can be said to be out
context in that they have no known close
tives, and their relationships to each
are unclear. More material must be
cted, especially from lower and middle
ocene rocks, before a valid family
gure can be erected for Oligocene
Dea
Except for the Squalodontidae, there-
we will consider the Oligocene Odon-
without assigning them to families.
Xenorophus
unique Oligocene odontocete is Xen-
Thus sloanii Kellogg (1923b) (Fig. 1a).
erophus was placed in the family Ag-
hidae by Miller (1923:40), and later
hors (Kellogg, 1928:32; Simpson, 1945:
Romer, 1966:392) have followed this
ment. The genus is founded on a par-
skull discovered during phosphate
ing operations at Woodstock near
leston, South Carolina, and sent to
logg by Earle Sloan. As reported by
ogg (1923b:2), Sloan stated that the
cimen came from 15 feet (4.6 m) be-
surface of the "Ashley-
the upper
per marl," the upper part of which
shley marl") Sloan regarded as "prob-
Oligocene" and the lower part
oper marl") as Upper Jackson (Eocene).
ugh now filled in, the marl pit which
mished the holotype of Xenorophus was
orted to be 76 feet (23.2 m) deep in
307
1917 (Cooke, 1936:83,85); thus, having
been found only 4.6 m below the top of the
marl, the specimen obviously came from
Sloan's "Ashley marl," which he surmised
to be of Oligocene age.
The age of the marl in the Charleston
area is of considerable importance in view
of the fact that these deposits yielded the
holotypes of both Xenorophus sloanii and
Agorophius pygmaeus (Müller, 1849) and,
as noted by Kellogg (1923a:27-28), pos-
sibly that of Archaeodelphis patrius Allen
(1921), a form also assigned to the Agoro-
phiidae by Miller (1923:40). Agorophius
has been suggested as "a somewhat dis-
tantly related precursor of the squalodonts"
(Kellogg, 1928:49) and Agorophius and
Archaeodelphis have been proposed as
"stages of development through which the
ancestors of some of the modern toothed
Cetacea might have passed" (Miller, 1923:
24-25). The Agorophiidae have even been
regarded as "the ancestral family" in the
Odontoceti (Rothausen, 1968:96; 1970:181-
183). These views are based upon the
cranial morphology of the forms involved,
but assignment of ancestral status to the
Agorophiidae hinges upon the age of the
deposits which produced the holotypes of
at least two of the three agorophiid taxa,
i.e. the Charleston marl beds.
Sloan's (1908:463–464) division of the
marl into two separately-named units
("Ashley marl" and "Cooper marl") was a
variation of previous concepts proposed by
Ruffin (1843), Tuomey (1848), Holmes
(1870) and Clark (1891), all of whom ex-
cept Ruffin (1843:7) referred these beds
to the Eocene along with the underlying
"Santee white limestone" of Lyell (1845:
434). Dall (1898:330,341) retained the
limestone in the Eocene but assigned the
marl to the lower Oligocene. Stephenson
(1914:85) applied the name "Cooper marl"
to the Charleston marl deposits and re-
garded them as a single formation "refer-
able to the uppermost Eocene or Oligo-
cene," as did Rogers (1914:186), who also
employed the name "Cooper marl."
Miller (1923:23) may have followed
[PAGE BREAK]
SYMPOSIUM: ADVANCES IN SYSTEMATICS OF MARINE MAMMALS
RAPRORE
PO
INTRODUCTION
CHARLES A. REPENNING
Geological Survey
U.S. Department of the Interior
345 Middlefield Road
Menlo Park, California 94025
On August 21, 1975, during the AIBS
eetings at Corvallis, Oregon, the Society
Systematic Zoology sponsored a sym-
ium on the Advances in Systematics of
rine Mammals. The topic was chosen
cause, since about 1960, this long-ne-
ected group of mammals has been the
ject of mushrooming interest: paleon-
logic, zoologic, and public. This interest
resulted in a great deal of new informa-
in with consequent improvement in our
derstanding of the history and evolution
these mammals. The symposium was
used primarily on paleontologic evi-
ce.
Twenty years ago, had a call been made
contributions to a symposium of this
only one paleontologist might have
ponded. This would have been Reming-
Kellogg who, for nearly 40 years, was
only authority in the United States on
fossil history of pinnipeds and cetaceans
one of the very few on fossil sirenians
the extinct desmostylians. Except for
ellogg, mammalian paleontology stopped
the shore of the sea.
Kellogg had many responsibilities, but
1962 he separated himself from the last
these by retiring from his position as
Director of the U.S. National Museum. He
en devoted his full time to his backlog
fossil marine mammals. By the time of
death seven years later he had pub-
hed nine major contributions to the sub-
Primarily these were concerned with
ecimens from the East Coast of the
aited States. At about this time, in 1961,
D. Mitchell began to publish a long
301
series of reports on fossil marine mammals
from the West Coast. In comparison to
earlier rates of publication, these of Kel-
logg and of Mitchell during the 1960's
amounted to a deluge. The interest they
generated in paleontologic circles was fur-
ther spurred by the growing interest in
marine mammals in zoologic circles. Thus
it was possible, in 1975, to devote an entire
symposium to the paleontologic evidence of
the systematics of marine mammals.
Fossil marine mammals are not so rare
as has been supposed and their phyloge-
netic histories are being reconstructed, con-
trary to the prior feeling that they were a
group whose record would always be so
poorly known that such understanding
would be impossible. Progress toward a
workable phylogenetic interpretation varies
with each type of marine mammal, how-
ever. This is partly a result of the fortunes
of discovery; partly a result of curatorial
abilities as illustrated by Sydney Ander-
son's axiom "It costs more to store a whale
than a mouse"; and partly due to differing
complexities in the evolution of particular
groups of marine mammals.
Interpretation of the phylogenetic history
of the cetaceans has been particularly de-
layed by Anderson's axiom and also by the
long and complex history of this group. The
cetaceans are unquestionably the most cos-
mopolitan of marine mammal groups and,
hence, are least susceptible to understand-
ing by study of fossils from a single oceanic
basin. However, the concept of faunal as-
semblages and of their succession through
time is now firmly fixed in the minds of
Systematic Zoology Val 25 De 1976 #4
Wash, P.C.
[PAGE BREAK]
302
paleocetologists, as is the awareness of
worldwide faunas. More than ever before
the fossil cetacean faunas of all oceanic
basins are being looked at as representing
the same historic events. Synonymies are
being recognized and histories that have
been documented in one part of the world
are being reinforced by what has been
learned in others. The possibilities of world-
wide correlation throughout most of the
Tertiary Period are becoming increasingly
interesting as the history of the Cetacea ap-
proaches its brink of discovery.
The sirenians present a similarly long
history but a far less cosmopolitan nature.
In addition, their evolution seems, at least
superficially, to be far less complex. This
lack of complexity is as troublesome as
great complexity. Slight differences that
have been used in the past as characters of
phylogenetic significance now seem to rep-
resent growth stages or ecologically dic-
tated specializations; other features have
remained distinctive, however, and the evo-
lutionary picture of the sirenians is now
much clearer. Restudy of forms described
as much as 100 years ago has been as en-
lightening as has the discovery of new fos-
sils. Possibly the most completely docu-
mented aspect of sirenian history is the
origin of the Steller Sea Cow, a genus
hardly brought to the attention of the sci-
entific community before it became extinct.
Although not a story of outstanding suc-
cess, its history has been one of remarkable
adaptation to environmental changes that
were overwhelming to many other organ-
isms; a history that is all the more spectacu-
lar in light of the striking lack of adapt-
ability shown by all other sirenians.
The extinct desmostylians, from present
information best thought of as four-legged
sirenians which lived in the North Pacific
something like 25 to 10 million years ago,
are little better off in our understanding
than they were ten years ago. Then the
only advancement in our awareness of the
group was the positive knowledge that they
really did have four legs. However, a re-
markably great number and variety of new
SYSTEMATIC ZOOLOGY
desmostylian fossils have recently begun to
be studied by Roy Reinhart of Miami Uni
versity, Ohio. Reinhart presented an intro-
duction to the material during the syn
posium but work on these fossils is
preliminary that he thought it impractical
to contribute to these published proceed
ings.
Due primarily to the pioneering efforts
of Edward D. Mitchell, and to the interest
he stimulated, much attention and much
understanding of the otarioid pinnipeds of
the North Pacific has developed. Of all
marine mammals, these pinnipeds are un
questionably the best known and their his
tory has been traced to their terrestrial
origins in fairly good, although not com-
pletely published, detail. Their implica
tions, biogeographic, biochronologic, eco-
logic, behavioral, and zoologic, are just
beginning to be recognized.
The history of the phocoid pinnipeds of
the North Atlantic has not fared so well
as has that of the otarioid pinnipeds. At
tention to their fossil record has lagged a
bit behind interest in the North Pacific
otarioids. Nevertheless, from the East
Coast of the United States to the Black Sea
to South Africa, with interesting hints from
other areas, concentrated effort has been
devoted to the discovery of their elution,
adaptations, and dispersal. Although major
gaps in fossil documentation still and will
continue to exist, understanding of the evo
lution of the phocoid pinnipeds is rapidly
forming a coherent picture.
Approaching the story from the direction
opposite to that of tracing their history
backward in time, a remarkably greater
understanding of probable pinniped origins
has come from the study of those terrestrial
carnivores which seem, in morphology and
historic position, to have been their prob
able ancestors. Although the early evolu
tion of the phocoids is still shrouded by
the lack of a fossil record, a much more
plausible understanding of their fissiped
ancestry has emerged from this study; the
essentially unquestionable origin of the
otarioids has been identified; and, at the
ODUCTION
time, a far more
anding of the evol
ivores has resulted.'
The symposium Adv
Marine Mammals w
current thinking. F
amaged to present the
rdless of their preli
ing the symposium
led to publish the
ngs in Systemati
ed that contributio
[PAGE BREAK]
394
The Fossil Book
telligent, nor especially novel; though they lived while archaic types died out,
they could not have survived really vigorous competition. But they did have
whatever was needed to produce descendants more able than themselves. They
therefore were first-rate ancestors, which means that we now must examine
their offspring, many generations removed.
MAMMALS TO THE SEA
We have found that reptiles, which evolved on land, soon sent invaders into
the water. Early mammals did the same, in the form of whalelike beasts known
as archaeocetes, zeuglodonts, or yoke-toothed whales.
The oldest known archaeocetes come from middle Eocene strata of north-
ern Africa and North America. Skulls suggest that their ancestors were
creodonts that presumably fed upon fish and spent more and more time in
the water. But the stages through which the creatures evolved are unknown,
for Eocene archaeocetes were fully aquatic. We see this in the pointed muz-
zle, elongate body, flipperlike forelegs, and hind legs so tiny that they were
hidden under flesh. The serrate cheek teeth had double roots, but teeth at the
front of the mouth were pointed or peglike. Nostrils had moved to the top of
the head, about halfway between the muzzle and eyes.
Archaeocetes culminated in Basilosaurus, also known as Zeuglodon ("yoke-
tooth"), whose remains are found in late Eocene deposits near the southern
Atlantic coast. Basilosaurus reached lengths of 55 to 70 feet, with a relatively
short head and a very long tail whose vertebrae were larger than those of the
short slim body. Tail vertebrae of this and other archaeocetes are frequently
found in the southeastern United States.
A few small, short-bodied archaeocetes lived on into Oligocene and earliest
Miocene times, but large ones died out at the end of the Eocene epoch. Their
place in the sea was taken by true whales, which probably were descended
from unspecialized archaeocetes. Early whales had short, stocky bodies, and
their nostrils formed a single blowhole that lay between or even behind the
eyes. Some genera possessed triangular teeth much like those of sharks, though
their bodies resembled porpoises. During Miocene and later times, however,
the group divided into true porpoises, beaked whales, sperm whales, and
whalebone whales. The first three of these retained their teeth and even in-
creased their number to several times the original forty-four. Whalebone
whales, however, became toothless and began to capture small animals in
plates of "whalebone"-really specialized skin-hanging down from the roof
of the mouth. Whalebone whales also have become the largest of all animals,
either living or extinct. The modern blue whale, for example, reaches lengths
exceeding 90 feet and weights greater than 120 tons. This living mammal,
therefore, is two to two and a half times as heavy as the largest dinosaurs.
MAKO
M
AKO SIKA-
old-time Sioux ref
canyons lying east
now applied to an
chiefly clay.
Clays of Mako S
is divided into a l
Brulé. They once f
isolated outcrops
katchewan. These
thirds of the Olig
grounds for remai
Because of its fi
was once suppose
now know that it
rolling hills and r
built during the
State Highway 22
called Independe
into its granite a
Grassy ground an
The climate of
and meandering
reedy lakes were
[PAGE BREAK]
340
Evolution of Vertebrates
The skull in the archaeocetes was somewhat more primitive than
the skull in later whales. The bones of the facial region were not
telescoped into the back of the skull, as they were in the more advanced
whales. There were forty-four teeth, as in primitive placentals, of which
the incisors and the canines were simple, sharp cones. The cheek teeth
were cusped, but with the cusps arranged in a single fore and aft
line, and coming to a high point in the middle. Teeth of this form
are seen in other mammals, especially certain seals that live on fish.
The nostrils in the archaeocetes were placed in a forward position,
not on top of the skull as in the later whales. In general this skull
appears as if it might have been derived from a creodont type, but
there is little beyond certain general resemblances to support such
a relationship. It must be emphasized that the ancestry of the Cetacea
is completely unknown at the present time.
ADAPTIVE RADIATION OF THE MODERN WHALES
During late Eocene or Oligocene times the modern whales arose as
descendants of the archaeocetes, and by Miocene times almost all the
families of modern whales had appeared. Two lines of cetacean evolu-
tion developed from the archaeocete stem. One of these was the
toothed whales or odontocetes; the other was the whalebone whales
or mysticetes.
Most of the modern whales are odontocetes. In late Oligocene times
there appeared some comparatively small odontocetes, designated
as the squalodonts, much like the modern porpoises in general appear-
ance, but characterized by cusped, triangular cheek teeth somewhat
similar to the teeth in the early archaeocetes. Evidently the squalo-
donts, of which the Miocene genus Prosqualodon is typical, were forms
intermediate between the archaeocetes and the modern types of whales.
In spite of their archaic teeth, the skull was highly advanced, with
the nostrils completely dorsal in position and the skull bones modified
accordingly. The squalodonts were important whales during the
Miocene epoch, but they did not survive long after the beginning of the
Pliocene epoch.
Their place in Pliocene, Pleistocene, and Recent times was taken
over by the small, toothed whales with which we are familiar, especially
the porpoises and dolphins, collectively known as delphinids. These are
compact, very swiftly swimming cetaceans that feed upon fish. The
teeth are greatly multiplied in number and have the form of simple
spikes. A common porpoise of Miocene times was Kentriodon; two of
the most widely spread of the modern porpoises are Delphinus, the
common dolphin, and Phocaena, the common porpoise of worldwide
Cetaceans
341
distribution. Related to these cetaceans are the fierce killer whales, the
so-called blackfish, and some of the river dolphins. Other small to
medium-sized odontocetes are the narwhals, the beaked whales, and
the platanistids that live in the Amazon and the Ganges rivers.
There was an early trend toward giantism in the toothed whales,
reaching its culmination in the physeterids or sperm whales. The
modern sperm whale or cachalot (Physeter) is a giant odontocete with
a great, square snout filled with gallons of sperm oil. Peglike teeth
are present on the lower jaw, and this whale feeds upon squids. In
the great days of whaling the sperm whale was much sought for its oil,
and incessant hunting has reduced it from its original vast numbers
to the limited herds of the present day. Moby Dick was a white sperm
whale.
The largest of the modern whales are the mysticetes or whalebone
whales, of much lesser variety than the toothed whales, but none
the less highly successful cetaceans. As mentioned above, these whales
feed upon plankton, and it may have been the abundance of their
food supply that led to the strong trend to giantism among these
largest of all vertebrates. The primitive mysticetes were the cetotheres.
in which the teeth had been lost and the skull had progressed toward
the high degree of modification that is so characteristic of the modern
whalebone whales. Mesocetus, of Miocene age, was typical of this group.
In the latter portion of Cenozoic times the evolution of the whale-
bone whales reached its ultimate stage with the development of the
skull into a highly arched structure for bearing large plates of baleen,
so that the eyes and braincase were limited to a very small posterior
region. Some modern representatives of this evolutionary trend are
the Greenland whales, the right whales (Balaena), the finbacks or
rorquals (Balaenoptera), and the titanic blue whales. These last may
reach lengths of one hundred feet and weights of one hundred and
fifty tons. We are wont to look back at some of the giant dinosaurs
with awe, yet such modern whales as the blue whale far exceed in size
the largest dinosaurs. These are the ultimate extreme of giantisin in
the evolution of animals.
WHALES AND MAN
A few hundred years ago whales roamed the seas in vast herds.
Then there was a demand for whale oil, and the large-scale systematic
hunting of whales began, reaching a high point in the last and present
centuries. Many species of large whales have been drastically reduced
in numbers. Whale oil is not used for lamps as it once was, but the
hunting of whales continues on a vast scale, for the oil is used for soaps,
MU ALISEMA Y
[PAGE BREAK]
I
E
er,
e
ed
esj
wh
on
re
rs
Australia. Rec. South Australia MUS., 11:35 wy
371.
HOMES, F. S. 1870. Phosphate rocks of South
Carolina and the "Great Carolina marl bed."
Charleston, S. C.
KELLOGG, R. 1923a. Description of two squa-
lodonts recently discovered in the Calvert Cliffs,
Maryland; and notes on the shark-toothed ceta-
ceans. Proc. U.S. Nat. Mus., 62, art. 16:1-69.
KELLOGG, R. 1923b. Description of an ap-
parently new toothed cetacean from South Caro-
lina. Smithson. Contrib. Knowl., 76, art. 7:1-7.
KELLOGG, R. 1924. Tertiary pelagic mammals of
eastern North America. Bull. Geol. Soc. Amer.
35:755-766.
KELLOGG, R. 1928. The history of whales-their
adaptation to life in the water. Quarterly Rev.
Biol., 3:29-76, 174–208.
KELLOGG, R. 1936. A review of the Archaeoceti.
Carnegie Inst. Washington, 1-366.
KEYES, I. W. 1973. Early Oligocene squalodont
cetacean from Oamaru, New Zealand. New
Zealand Jour. Marine and Freshwater Research,
7(4):381-390.
LYDEKKER, R.
CUATRO
Tertiary cetacean from Oregon. Northwest Sci.
ence, 49:174-181.
RABEDER, G. 1975. Die Wirbeltierreste (excl.
Pisces) aus dem Egerien von Österreich: p.
427-453 in OM Egerien, Die Egerer, Pouz
dřaner, Puchkirchener Schichtengruppe und die
Bretkaer Formation: Chronostratigrape und
Neostratotypen, Miozän der Zentral
Par-
atethys, Bd. V. VEDA, Verlag der Slow kischen
Akad. Wiss., Bratislava.
ROGERS, G. S. 1914. The phosphate deposits of
South Carolina. U.S. Geol. Survey Bull. 580,
pt. 1:183-220.
ROMER, A. S. 1966. Vertebrate paleontology.
3rd ed., Univ. Chicago Press, 1-468.
ROTHAUSEN, K. 1958. Marine Vertebraten
(Odontaspidae, Lamnidae, Sparidae, Der-
mochelyidae, Squalodontidae) im oberoligo-
zänen Meeressand von Süchteln und Dusseldorf.
Fortschr. Geol. Rheinld. u. Westfall., 1:363–381.
ROTHAUSEN, K. 1968. Die systematise: Stel-
lung der europäischen Squalodontidae Odont-
oceti, Mamm.). Paläont. Zeit., 42:80-01.
1892.
ROTHAUSEN,
On zeuglodont and other
cetacean remains from the Tertiary of the
Caucasus. Proc. Zool. Soc. London for 1892,
part 4:558-564.
LYELL, C. 1845. Observations on the white
limestone and other Eocene or older Tertiary
formations of Virginia, South Carolina, and
Georgia. Geol. Soc. London Quart. Jour.,
1:429-442.
MCKAY, A. 1882. On the younger deposits of
Wharekauri Basin and the lower Waitaki Val-
ley. Repts. Geol. Explor. 1881, Colonial Mus.
and Geol. Surv. Dept., Wellington, New
Zealand: 98–106.
MALDE, H. E. 1959. Geology of the Charleston
phosphate area, South Carolina. U.S. Geol. Sur-
vey Bull. 1079:1-105.
MARPLES, B. J. 1956. Cetotheres (Cetacea)
from the Oligocene of New Zealand. Proc. Zool.
Soc. London, 126:565-580.
MCHEDLIDZE, G. A. 1970. Nekotorye obshchie
cherty istorii kitoobraznykh. Ch. 1. "Mets-
niereba," Tbilisi: 1-111. Transl. by Dorothy
B. Vitaliano, U.S. Geological Survey TR 71-14.
MILLER, G. S. 1923. The telescoping of the
cetacean skull. Smithsonian Misc. Collections,
76(5):1-68.
NORRIS, K. S. 1968. The evolution of acoustic
mechanisms in odontocete cetaceans. In Drake,
E. T., ed., Evolution and Environment, Yale
Univ. Press, 297-324.
NORRIS, K. S. 1975. Cetacean biosonar: a two
part review. Part 1. Anatomical and behavioral
studies. In Malins, D. C. and Sargent, J. R.,
eds., Biochemical and Biophysical perspectives
in Marine Biology, Academic Press, 215-236.
K. 1970. Marine Reptilia and
Mammalia and the problem of the Oligocene.
Miocene boundary. Giornale di Geologia (2)
35, fasc. 1:181-189.
ROTHAUSEN, K. 1971. Cetotheriopsis tobieni n.
sp., der erste paläogene Bartenwal (Ceto-
theriidae, Mysticeti, Mamm.) nördlich des
Tethysraumes. Abh. hess. L.-Amt Bodenforsch.
60:131-148.
n the
RUFFIN, E. 1843. Report of the commencement
and progress of the agricultural survey of South
Carolina for 1843. Columbia, S. C., p.
RUSSELL, L. S. 1968. A new cetacean
Oligocene Sooke Formation of Vi conver
Island, British Columbia. Canadian J. Earth
Sci., 5:929-933.
SIMPSON, G. G. 1945. The principles of clas
sification and a classification of mammals. Bull.
Amer. Mus. Nat. Hist., 85:1-350.
SLOAN, E.
odified their diet in this way, two sina
ken up a very different mode of life-th
ve been accused of this and other crime
roof of blood-sucking among the Chiro
erformed. Chisel-like upper incisor teet
with little pain or inconvenience to the
vound.
s been limited to forms familiar in north
elatives. There exists, however, a secon
confined to the Old World tropics-th
form a distinct suborder, termed th
ontrast to the familiar little members o
well deserved, for they are of relatively
ving spread. The popular name is like
a well-developed, foxlike muzzle, with
y of the members of the other group
although these animals are able flyers
system of their smaller cousins. Such
s group, for their dict consists of fruits
ans of t.avel between the trees, where
e swarms-and the forests or orchards
side branch, and the stem of the order
y be sure that the bats arose from the
ch had a similar diet. Fortune has
pre-
the Eocene in which the flight struc-
1908. Catalogue of the mineral lopped as today. Farther back, however,
calities of South Carolina. S. C. Geol. Survey.
ser. 4, Bull. 2:1-505.
STEPHENSON, L. W. 1914. A deep well at
Charleston, S. C. U.S. Geol. Survey Prof. Paper
90:69-94.
THENIUS, E.
1969. Phylogenie der Mamalia-
Walter de Gruyter & Co., Berlin: 1-72
TRUE, F. W. 1907. Remarks on the type of the
fossil cetacean Agorophius pygmaeus (Müller).
Smithsonian Inst. No. 1694:1-8.
TUOMEY, M. 1847. Notice on the discovery of a
cranium of the zeuglodon. Proc. Acad. Nat. Sci.
Phila., 3:151-153.
1-
TUOMEY, M. 1848. Report on the geology of
South Carolina. Columbia, A. S. Johnson.
293.
prins which must have existed at the
tor, the "flying lemur" of the East
age in the development of flight. As
remarked, the name is hardly appro-
hor does it fly. It does, however, have
ick from its arms by means of which
nce. Probably the earliest bats began
PORPOISES
rpoises constitute the largest and
at has turned to an aquatic life and
COL
[PAGE BREAK]
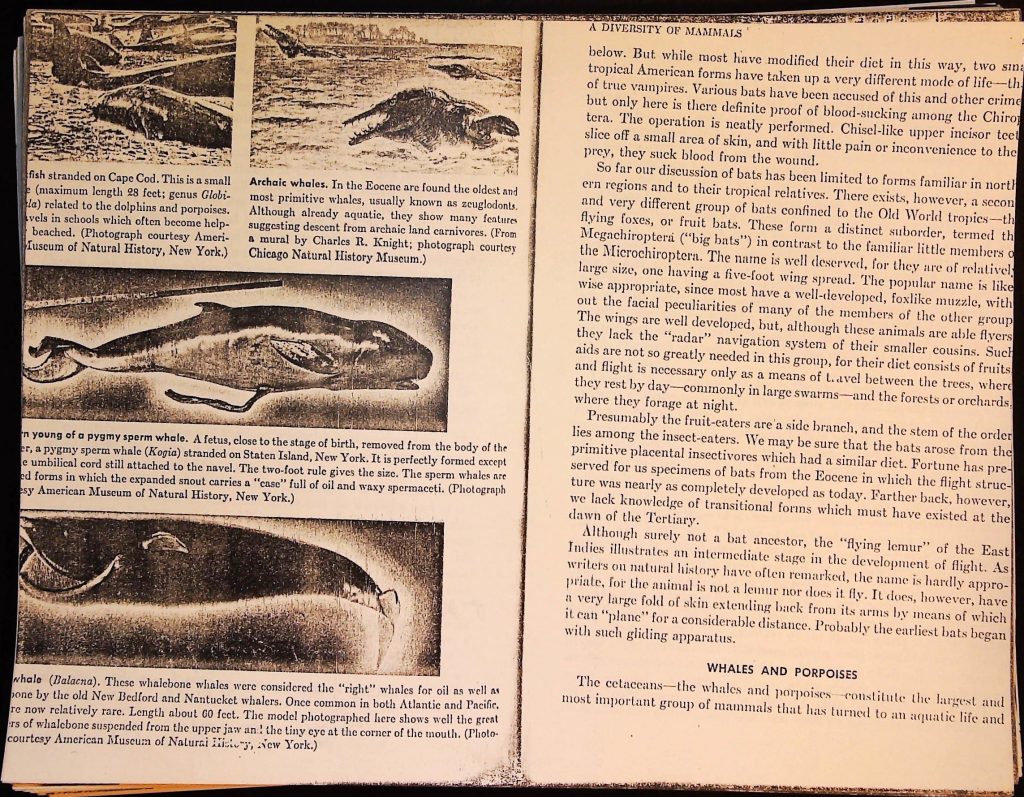
fish stranded on Cape Cod. This is a small
e (maximum length 28 feet; genus Globi-
la) related to the dolphins and porpoises.
vels in schools which often become help-
beached. (Photograph courtesy Ameri-
Iuscum of Natural History, New York.)
Archaic whales. In the Eocene are found the oldest and
most primitive whales, usually known as zeuglodonts.
Although already aquatic, they show many features
suggesting descent from archaic land carnivores. (From
a mural by Charles R. Knight; photograph courtesy
Chicago Natural History Museum.)
n young of a pygmy sperm whale. A fetus, close to the stage of birth, removed from the body of the
er, a pygmy sperm whale (Kogia) stranded on Staten Island, New York. It is perfectly formed except
e umbilical cord still attached to the navel. The two-foot rule gives the size. The sperm whales are
ed forms in which the expanded snout carries a "case" full of oil and waxy spermaceti. (Photograph
esy American Museum of Natural History, New York.)
whale (Balacna). These whalebone whales were considered the "right" whales for oil as well as
one by the old New Bedford and Nantucket whalers. Once common in both Atlantic and Pacific.
re now relatively rare. Length about 60 feet. The model photographed here shows well the great
rs of whalebone suspended from the upper jaw an! the tiny eye at the corner of the mouth. (Photo
courtesy American Museum of Natural History, New York.)
A DIVERSITY OF MAMMALS
below. But while most have modified their diet in this way, two sina
tropical American forms have taken up a very different mode of life-th
of true vampires. Various bats have been accused of this and other crime
but only here is there definite proof of blood-sucking among the Chirop
tera. The operation is neatly performed. Chisel-like upper incisor teet
slice off a small area of skin, and with little pain or inconvenience to the
prey, they suck blood from the wound.
So far our discussion of bats has been limited to forms familiar in north
ern regions and to their tropical relatives. There exists, however, a secon
and very different group of bats confined to the Old World tropics-th
flying foxes, or fruit bats. These form a distinct suborder, termed th
Megachiroptera ("big bats") in contrast to the familiar little members o
the Microchiroptera. The name is well deserved, for they are of relativel
large size, one having a five-foot wing spread. The popular name is like
wise appropriate, since most have a well-developed, foxlike muzzle, with
out the facial peculiarities of many of the members of the other group
The wings are well developed, but, although these animals are able flyers
they lack the "radar" navigation system of their smaller cousins. Such
aids are not so greatly needed in this group, for their diet consists of fruits
and flight is necessary only as a means of t.avel between the trees, where
where they forage at night.
they rest by day-commonly in large swarms-and the forests or orchards.
Presumably the fruit-eaters are a side branch, and the stem of the order
lies among the insect-eaters. We may be sure that the bats arose from the
primitive placental insectivores which had a similar diet. Fortune has pre-
served for us specimens of bats from the Eocene in which the flight struc-
ture was nearly as completely developed as today. Farther back, however,
dawn of the Tertiary.
we lack knowledge of transitional forins which must have existed at the
Although surely not a bat ancestor, the "flying lemur" of the East
Indies illustrates an intermediate stage in the development of flight. As
writers on natural history have often remarked, the name is hardly appro-
priate, for the animal is not a lemur nor does it fly. It does, however, have
a very large fold of skin extending back from its arms by means of which
with such gliding apparatus.
it can "plane" for a considerable distance. Probably the earliest bats began
WHALES AND PORPOISES
The cetaceans the whales and porpoises constitute the largest and
most important group of mammals that has turned to an aquatic life and
[PAGE BREAK]
300
THE VERTEBRATE STORY
to the common stem of the group. Typically there is a good battery of
peglike teeth. Porpoise-like cetaceans became common in mid-Tertiary
times; in their very long snouts and in some further technical respects, the
river dolphins of today are regarded as closer to these older types than are
their marine cousins. Such fresh-water forms are found in the Amazon and
La Plata rivers in South America, in the Ganges and Indus rivers of
India, and in a single lake far inland in China. As might reasonably be
expected from the relatively small bodies of water in which they navigate,
these dolphins are small, from five to eight feet in total length. The Indian
dolphin is quite aberrant in that it is blind, probing the mud of the river
bottoms for concealed fishes and fresh-water shrimps.
Among the numerous marine members of this group of smaller cetaceans
there is considerable variation. The term "porpoise" is generally applied to
the smaller members of the family which are short-snouted. The familiar
and playful Atlantic porpoise never exceeds six feet in length, and there
are a number of other species of similar size. The term "dolphin" properly
belongs to larger forms, with pronounced noses. For example, along the
Atlantic coast of North America the animal frequently called a "porpoise"
is actually the common bottle-nosed dolphin, a larger animal (up to twelve
feet in length). The dolphin of the ancients is a type with worldwide dis-
tribution but is especially abundant in the Mediterranean. It averages
eight feet or so in adult length and has a prominent beak.
Two interesting members of the family inhabit cold northern seas. The
"white whale," or beluga, is unique in its pure-white coloration, but since
it seldom reaches a length of more than fourteen to sixteen feet, it hardly
deserves to be called a whale. Schools of this gregarious cetacean may be
found as far south as the St. Lawrence River and the northern part of the
Norway coast. Related and similar in many ways is the narwhal, notable
for the presence-in the male only-of a greatly elongated and spirally
twisted tusk, formed by the elongation of an incisor tooth. Possibly this
structure may be used for dueling purposes by males in the mating season.
(This is pure speculation, but no other reasonable explanation has ever
been given.)
Two final members of the dolphin family are often called "whales" with
greater justice, if size be the criterion. The blackfish, or pilot whale, a
feeder on cuttlefish, is found in large herds in the North Atlantic; it is
black in color, as its name suggests, and has a rounded, swollen forehead;
it may grow to as much as twenty-eight feet. Similar in size, but very dif-
ferent in other respects, is the "killer whale"; packs of this black and
A DIVERSITY OF MAMMALS
white spotted form may be found in every ocean region. This is an aggre
sive and vicious animal; it feeds to some extent on salmon and other fishe
but sea birds and seals are its favorite food; it preys upon other membe
of the porpoise-dolphin group; still further, a killer pack does not hesita
to attack even the very largest of whales, many times their own size.
The remaining toothed cetaceans are unquestionably whales. They a
few in numbers of genera and species. Three types may be distinguished-
the beaked, bottle-nosed, and sperm whales. The beaked whales get the
name from the presence of a distinct projecting beak comparable to that
a dolphin. The greatest length they attain is about twenty-six feet, an
some species are much smaller. Although technically toothed whales, the
are, in fact, almost toothless, for the only teeth in the adult are usually
pair in the lower jaw, and even these may be much reduced. These form
are of little commercial importance. The bottle-nosed whales, somewha
larger in size, also have beaks, but this feature is less prominent because o
the presence above it (as in the similarly named dolphin) of a bulging fore
whale relatives.
head. The teeth are reduced in a fashion similar to that of their beaked
Largest of all toothed whales is the great sperm whale, or cachalot
Reports of individuals eighty feet or more in length appear to be on a pa
with fishermen's stories, but authenticated measurements of sixty feet o
so testify to animals of considerable bulk. Curiously, however, this anima
has a tiny relative, the pygmy sperm whale, which resembles it in every
feature except size, for this little animal when mature
a dozen feet long.
seldom more than
In body proportions the great sperm whale is very different from all
other cetaceans because of its enormous head, bounded below by a short
and slender lower jaw. The size of the head has nothing to do with the
skeleton; the front and upper parts of this massive structure are occupied
by tissues surrounding a great reservoir of oil, called "spermaceti"-as
much as fifteen barrels of it in a large individual. This is presumably a
valuable store of reserve food for the whale--and made the sperm whale a
valuable catch for the whalers of the pre-petroleum days. Apart from the
spermacetic reservoir, the structure of the sperm whale is fairly similar
to that of the beaked and bottle-nosed forms, and even this reservoir has a
smaller counterpart in the latter form. As in those whales, the dentition is
much reduced, although the sperm whale retains a row of small lower
teeth. Tooth reduction may be related to diet, for the sperm whale, like
its beaked and bottle-nosed relatives, feeds on squids and cuttlefish.
[PAGE BREAK]
302
THE VERTEBRATE STORY
The sperm whale is also the source of ambergris, a lightweight material
offensive in odor when fresh but valuable as a perfume base; masses of it
are frequently found floating in the sea or cast up on the shore. The mate-
rial originates in the whale's intestine; horny cuttlefish beaks are fre-
quently found embedded in it, and it has been suggested that they form
the nucleus for its formation. (Note to fortune-seekers: specimens of sup-
posed ambergris brought to museums for identification usually turn out to
be sewage!)
Whalebone whales.-Sharply marked off from the toothed forms are
the whalebone whales. Here teeth have been lost entirely. The huge mouth
The right whale (abore) and sperm
whale (below); both have large
heads but that of the right whale is
mainly occupied by the large
mouth, filled with whalebone
plates, while that of the sperm
whale consists mainly of the great
spermaceti sac, lying above the
upper jaws.
is filled with a series of sheets of whalebone,
fringed with hairs at the edge and hanging
down from the roof of the mouth in parallel
rows like the leaves of a book. This whale-
bone is composed of hardened horny skin.
In the roof of the mouth of the dog, for ex-
ample, may be seen crosswise ridges of skin;
whalebone is formed by the elaboration of
such a series of skin ridges.
These whalebone whales live upon plank-
ton, small animal organisms found in sea
water, particularly a type of tiny shrimp:
water, passing through the mouth, is strained
through the whalebone filter, and the edible
material deposited is licked off by the tongue.
It would be impossible for this type of whale
to swallow any large object (such as Jonah!);
although the mouth opening may be huge, the gullet does not exceed nine
inches in diameter. It is paradoxical that these eaters of tiny food par-
ticles should include in their number the largest animals of any kind that
have ever existed.
Best known of whalebone whales are the "right whales." These whales
get their name simply from the fact that in early days this was the right
kind of whale to hunt because, in addition to a good yield of blubber, its
whalebone was of excellent quality and, unlike some other whales of the
group, the animals would not sink when killed. Now rare in most areas, the
right whales once ranged the world over. There are a number of species; a
pygmy member of the group is only twenty feet long, but typical right
whales generally run to about sixty feet in length as adults. The head is
wh
qua
Hist
AL
the
the
[PAGE BREAK]
A DIVERSITY OF MAMMALS
303
large and blunt, with about the proportions of that of a sperm whale. But
on even superficial inspection, it can be seen that it is built in very different
fashion. In the sperm whale the head proper is enormous, the lower jaw
short and slender. Here the head appears to consist mainly of lower jaws,
the line of the long mouth-opening curving up, back, and then down, end-
ing close to the small eye. This build of the jaws is necessary to give a
cover over the enormous series of plates of whalebone, several hundred in
number, hanging down from the roof of the mouth.
The rorquals, or fin whales, are today the most important animals in the
whale fisheries; they were formerly neglected because the whalebone was
of poor quality and smaller in amount, and-more important in earlier
times because the carcass was hard to deal with since it tended to sink
to the bottom. The head is broad but much more slender in side view than
that of a right whale due to the lesser depth of the whalebone "battery";
the flippers, too, are more slender and tapering than in the right whales.
The "lesser" rorqual runs to but thirty feet in length; on the other hand,
the great blue whale, or sulfur-bottom, a member of this group, is the
largest animal in the world, reaching a maximum length of nearly one
hundred feet and (judging by the known weight of smaller individuals) a
weight close to two hundred tons. Related to the rorquals is the humpback
whale, a fifty-foot animal with a stout build and very long flippers. Quite
distinct from either right whales or rorquals is the "California" gray
whale, a relatively slender-headed animal that never reaches more than
forty-five feet in length. It inhabits the North Pacific, generally spending
the summer in the Arctic and descending along the American and Siberian
coasts in the winter.
Archaic whales.-Little is known of either the toothed or whalebone
whale groups before the Miocene epoch. In the Eocene, however, archaic
whales, the zeuglodonts, were already common in the seas. These were, in
some cases, of considerable size (seventy feet is the apparent maximum),
with a body comparatively longer but sliminer than later whales; the
proportions were those which modern imagination ascribes to the sea
serpent. These forins were considerably closer to their land ancestors than
the typical whales. In the skull, for example, the nostrils were still near the
front of the head, and the teeth were very much like those of primitive
flesh-eating mammals. Not improbably the whales came from early creo-
dont carnivores which gradually took up a fish-eating existence in the way
that otters and seals later did; but we have no fossil record of the early
stages in the transition from land to water.
[PAGE BREAK]
e),
that
Natural
the whole &
THE CETACEAN DIVERSITY
hands. The upper jaw-bones, the maxillae, are pulled forward to make
a beak or rostrum, and at the same time the premaxillae are extended
backwards from the tip of the rostrum to push the nostrils back towards
the frontal bone. As this happens the nasal bones, which roof the nasal
passage, are pushed back towards the frontal, and greatly reduced in
size, so that in some species they are represented only by a small
tubercle on that bone. While these bones of the fore part of the skull are
extending backwards, the supraoccipital bone at the back of the skull is
drawn upwards and forwards to cover the brain and form the posterior
part of the roof of the brain case. So extensive is its proliferation that it
pushes the parietals, which in most mammals form the sides and roof of
the cranium, away to right and left so that they do not make a suture in
the mid-line. The frontal is thereby much reduced in size so that it takes
the form of a fillet on the forehead, expanded laterally over the orbits.
As these changes are made numerous modifications of the other
bones of the skull are necessary. The ethmoid thus forms the greater
part of the front of the cranium, supported and partly overlain by the
unusually large expanded pterygoids, which, with the vomer, extend
forwards, largely usurping the place of the palatine. At the same time
the jugal arch is much reduced, often to a slender bar below the orbit,
and in some species is completely lost; but the squamosal remains
relatively large. The ear bones, the tympanic and periotic, are only
loosely articulated with the rest of the skull.
The combined result of these modifications is that the brain case is
short but high and wide, the nasal passage lies approximately vertically
in front of it, and a comparatively long rostrum extends approximately
horizontally before. This basic cetacean pattern is modified in various
directions in the different suborders and families.
The order Cetacea is divided into three suborders; the Archaeoceti,
all of which are extinct and known only from fossil remains, the
Odontoceti or toothed whales, containing the majority of the living
species forming the order; and the Mysticeti, the whalebone or baleen
whales.
The Archaeoceti, as their name implies, are the oldest group and
consequently the most primitive in their structure. They flourished
during the Eocene epoch but most of them were extinct before the end
of the Oligocene, only a few small species surviving into the early
Miocene. The most primitive species, such as Protocetus, were elon-
gated aquatic animals with reduced hind limbs and long snouts. The
25
[PAGE BREAK]
THE CETACEAN DIVERSITY
Suborder
ARCHAEOCETI†
Families
Protocetidae†
Dorudontidae*
Basilosauridaet
(=Zeuglodontidae†)
Order CETACEA
Th
the s
striki
Suborder
Suborder
inde
ODONTOCETI
MYSTICETI
their
horiz
Families
Families
body
Agorophiidae†
Cetotheriidaet
verte
Squalodontidaet
Rachianectidae
para
Platanistidae
Ziphiidae
Balaenopteridae
Balaenidae
close
This
Physeteridae
mov
Eurhinodelphidae*
Hemisyntrachelidaet
Acrodelphidae†
Monodontodidae
Delphinidae
Phocaenidae
mas
poss
wat
Cru
hav
the
tail-
† extinct
Figure 2. The classification of the Cetacea.
bones of the skull are all arranged in the typical mammalian pattern
without the distortion or 'telescoping' shown by modern Cetacea. The
only sign of events to come is the position of the nostrils on the top of the
snout or rostrum some way back from the tip. The teeth, too, are
completely different from those of modern whales for they are hetero-
dont, and distinguishable as incisors, canines and grinders, the last
sectorial rather than molar, probably in correlation with a diet of fish.
The dentition and general form of the skull show that the early Cetacea
were probably descended from a common stock with the primitive
Creodonta, which also gave rise to the Artiodactyla as well as the
Carnivora.
The most striking and characteristic family of the Archaeoceti is the
Basilosauridae or Zeuglodontidae, members of which were abundant in
the seas of the upper Eocene. They were large animals, some species
reaching a length of 70 feet, and having skulls 5 feet long. Unlike the
modern whales they were not fusiform in body shape but snake- or
eel-like; elongated and cylindrical. The neck was short but the rest of
the vertebral column very long; the fore-limbs were short paddles and
the hind limbs had been lost.
26
on
told
the
Ba
Ha
wh
IOQ
hu
an
Sd
fr
Z
Pl
St
as
[PAGE BREAK]
n
e
e
e
t
a
e
e
E
THE CETACEAN DIVERSITY
The form of the teeth suggests that these creatures fed upon fish, but
the snaky shape of the body suggests that they may have differed
strikingly from the rest of the Cetacea in the shape of their tail flukes, if
indeed they had any. The snake- or eel-like body leads to speculation on
their way of swimming. Did they swim by vertical oscillations of
horizontal tail flukes, or did they swim by lateral flexures of the whole
body, as in the animals they resemble, albeit on an enormous scale? The
vertebrae each bear dorsal and lateral processes which are com-
paratively small so that there was no possibility of their articulating
closely with each other or being tightly bound together with ligaments.
This gave the vertebral column a degree of flexibility and freedom of
movement that is denied to the modern Cetacea which have much more
massive processes articulating to form a more rigid spine. It is therefore
possible that the basilosaurs lived like eels in comparatively shallow
water, snaking their way over the bottom to snap up the fish and
Crustacea of the benthic fauna. If this were so, they probably did not
have horizontal tail flukes, nor indeed flukes of any sort: the hind end of
the body and the tail were no doubt laterally compressed, as is the
tail-stock of all modern cetaceans, and this would give all the purchase
on the water needed in eel-like swimming.
The long snake-like form of the zeuglodonts misled the palaeon-
tologists who examined the first specimens discovered into thinking
they were dealing with the remains of an extinct reptile, hence the name
Basilosaurus bestowed upon them in 1834 by the American geologist
Harlan but altered in 1839 to Zeuglodon by the English anatomist Owen,
who based his name on the character of the molar teeth, which have two
roots joined by a blade-like crown.
In the second quarter of the nineteenth century an American fossil
hunter, Dr Albert C. Koch, collected a large number of fossil vertebrae
and other bones of archaeocetes from the Eocene beds in Alabama and
South Carolina, where they are abundant. Although the vertebrae came
from several individuals Dr Koch strung them together to make the
skeleton of a supposed animal 114 feet long, which he exhibited as the
skeleton of a sea-serpent in a hall on Broadway, New York, in 1845.
Zoologists were quick to denounce the imposture but nevertheless the
public, as always eager for marvels, flocked to see it so that the self-
styled doctor made good thing out of his humbug. He even went so far
as to give the creature a scientific name, Hydrarchos sillimani, in honour
of Professor Benjamin Silliman, a genuine scientist who for many years
27
[PAGE BREAK]
HOW
C
dn
HOU
ed t
ons
0-A
HO
M
an
Bɔ1
Te
19
SIE
ea
30
J
00
150
21
чо
THE CETACEAN DIVERSITY
edited the American Journal of Science and Arts, also known as Silliman's
Journal of Science. The doctor, whose own name is not inappropriate,
seems to have had a cynical sense of humour in selecting the name of the
well-known professor for his fabrication. Now we have something even
sillier - Nessiteras rhombopteryx, P. Scott 1975, the non-animal of Loch
Ness.
Among the Archaeoceti the Protocetidae may represent the stock that
gave rise to both groups of living whales, whereas the Basilosauridae,
although having skulls and dentition that were primitive as compared
with those of modern whales, were highly evolved and specialized in
their post-cranial structure. They can in no way be regarded as the
progenitors of the order Cetacea; they are no more than an aberrant
branch from the common stock of origin.
Most of the members of the Odontoceti are comparatively small
porpoises and dolphins, though some, such as the Beaked whales and
the Killer whale, reach a length of 30 feet and one, the Sperm whale,
reaches 60 feet or more. In most of the odontocetes the jaws are
prolonged more or less as a beak-like snout behind which the forehead
rises in a rounded curve that is very prominent in some species in which
it is called the 'melon': melon in colloquial French means a bowler hat.
Even in those species that lack a beak the rostral part of the skull is
prolonged forwards so that if the flesh is removed the skull shows the
same general form as that of the beaked species.
The Odontoceti differ from the other suborders in the arrangement
of their nasal passages and blow-holes; they differ indeed from all other
mammals in having only a single nostril. The nasal passages are separate
at the base of the skull, as is usual, but in their passage to the surface
they join close below it to form a single opening or, in extreme cases, one
is functionally suppressed leaving the other as the sole breathing tube.
The blow-hole is typically a crescentic slit, with the horns directed
forward on the summit of the head; it is closed by a fatty and fibrous pad
or plug that lies between the horns. When the animal breathes it thus
has to open the blow-hole by muscular effort; when it dives the pressure
of the water keeps the hole sealed. There are, in addition, various side
branches and valvular arrangements inside the blow-hole which will be
discussed below when dealing with the respiration of the Cetacea
(Chapter 5).
The skull bones of the nasal region, especially the premaxillae and the
nasal bones, are often asymmetrical in their size, shape and position.
28
f
[PAGE BREAK]
THE AGE OF MAMMALS
Figure 17. Archaic whale, Prozeuglodon of the late Eocene, was as large as a
porpoise but still very primitive in many respects. Its serrated cheek teeth may
suggest that whale ancestry lay within Creodonta or Carnivora.
Moeritherium shows some resemblance to the Sirenia or sea-cows, which
in fact are regarded as an offshoot of the same stock as the proboscideans,
and thus also should be of African origin. Sea-cows are, in fact, found for the
first time in these Egyptian deposits, and in strata of about the same age
in Jamaica; so the Sirenia were widespread at this early date. The early
sirenians show the characters of the order in an incipient stage
peculiar massiveness of the bone structure, the downturning of the snout,
and the gradual reduction of the teeth.
the
Of greater importance are the whales, which make their appearance in
the middle Eocene of Egypt; whether this really proves an African origin
is doubtful, however, for at least one specimen has also been found in the
mid-Eocene of North America, and one fragment from the lower Eocene
of England proves the presence of whales in the Atlantic at that early date.
In the late Eocene, whales had invaded the seas in large numbers and great
variety and are found in marine deposits of North America, Europe, and
Africa.
The earliest whales, or Archaeoceti, show a rapid adaptation to a com-
pletely aquatic life. The hind legs were reduced to vestiges not projecting
from the body, just as in modern whales. The largest of the early whales,
the zeuglodonts of the late Eocene (Basilosaurus, etc.), were somewhat
serpent-like creatures up to fifty-five feet in length and must have re-
sembled certain sea-going lizards of the Cretaceous very closely. (A
famous maker of fake exhibitions in the nineteenth century combined the
vertebrae of several individuals to create a monster a hundred metres long.)
The serpentine body of the archaeocetes is more primitive than the com-
pact body of a modern cretacean. The head was comparatively small and
separated from the body by a distinct neck, while the large head of a
modern whale is joined to the body without an externally visible neck.
The head of the archaeocetes is not unlike that in certain creodonts, but
80
the snout was elon
head. The front te
ful, shearing struct
for the middle Eoc
some member of t
the seals were to e
Eocene history
complicated one, a
teen million years
Asia will probabl
Eocene date are al
faunas. In Africa
still provide infor
Tertiary. At prese
from the Eocene
Again, when w
Eocene, the recor
rises from forty-o
Of course, it sho
really due to inc
Still, the main pa
If the Paleocer
as an epoch of co
dominant group
Table 4. The Eoc
Epoch
G
Oligocene
Fayu
Eocene
Paleocene
F
Mok
[PAGE BREAK]
A
ich
ns,
the
age
rly
the
but,
gin
the
ene
ate.
[eat
and
THE EOCENE: EPOCH OF CONSOLIDATION
the snout was elongate and the nostrils were half-way up to the top of the
head. The front teeth were simple cones, but the cheek teeth were power-
ful, shearing structures not unlike those in creodonts; this is especially true
for the middle Eocene archaeocetes and suggests that the whales arose from
some member of the Carnivora or Creodonta which lived on fish, just as
the seals were to evolve much later from swimming carnivores.
Eocene history in both North America and Europe is a very long and
complicated one, as may be seen from Table 4. It spans eighteen or nine-
teen million years and is subdivided into four or five ages. The blanks in
Asia will probably be filled in before long; faunas of early and middle
Eocene date are already known, though not so well as the great late Eocene
faunas. In Africa the situation is less promising, but diligent search may
still provide information on the land faunas of this continent in the early
Tertiary. At present it comes into focus with the Fayum and the transition
from the Eocene to the Oligocene.
Again, when we look at the familial diversity of the mammals in the
Eocene, the record is one of great increase. The number of families known
rises from forty-one in the late Paleocene to ninety-one in the late Eocene.
Of course, it should be remembered that part of the seeming increase is
really due to incompleteness of the Paleocene and early Eocene record.
Still, the main part of the increase is probably real.
If the Paleocene was an epoch of conquest, the Eocene may be regarded
as an epoch of consolidation. The mammals had definitely emerged as the
dominant group of land animals and were now crowning their victory by
Table 4. The Eocene of the World Continent
m-
ung
Epoch
Africa
(faunas)
Asia
(faunas)
North
America
(ages)
Europe
(ages)
Date
(Million
years)
les,
bat
Oligocene
(Mongolia) Chadronian
Sannoisian
37
Fayum
re-
Irdin Manha Duchesnean
Pondaung
Ludian
(A
the
ag.)
40
Mokattam
Eocene
m-
Uintan
Bartonian
45
Bridgerian
Lutetian
49
and
Ulan Bulak
Ypresian
of a
Wasatchian
Sparnacian
55
eck.
Paleocene
Gashato
Clarkforkian
but
F
81
[PAGE BREAK]
THE AGE OF MAMMALS
Figure 17. Archaic whale, Prozeuglodon of the late Eocene, was as large as a
porpoise but still very primitive in many respects. Its serrated check teeth may
suggest that whale ancestry lay within Creodonta or Carnivora.
Moeritherium shows some resemblance to the Sirenia or sea-cows, which
in fact are regarded as an offshoot of the same stock as the proboscideans,
and thus also should be of African origin. Sea-cows are, in fact, found for the
first time in these Egyptian deposits, and in strata of about the same age
in Jamaica; so the Sirenia were widespread at this early date. The early
sirenians show the characters of the order in an incipient stage.
peculiar massiveness of the bone structure, the downturning of the snout,
and the gradual reduction of the teeth.
the
Of greater importance are the whales, which make their appearance in
the middle Eocene of Egypt; whether this really proves an African origin
is doubtful, however, for at least one specimen has also been found in the
mid-Eocene of North America, and one fragment from the lower Eocene
of England proves the presence of whales in the Atlantic at that early date.
In the late Eocene, whales had invaded the seas in large numbers and great
variety and are found in marine deposits of North America, Europe, and
Africa.
The earliest whales, or Archaeoceti, show a rapid adaptation to a com-
pletely aquatic life. The hind legs were reduced to vestiges not projecting
from the body, just as in modern whales. The largest of the early whales,
the zeuglodonts of the late Eocene (Basilosaurus, etc.), were somewhat
serpent-like creatures up to fifty-five feet in length and must have re-
sembled certain sea-going lizards of the Cretaceous very closely. (A
famous maker of fake exhibitions in the nineteenth century combined the
vertebrae of several individuals to create a monster a hundred metres long.)
The serpentine body of the archaeocetes more primitive than the com-
pact body of a modern cretacean. The head was comparatively small and
separated from the body by a distinct neck, while the large head of a
modern whale is joined to the body without an externally visible neck.
The head of the archaeocetes is not unlike that in certain creodonts, but
80
the snout was elongate
head. The front teeth
ful, shearing structures
for the middle Eocene
some member of the
the seals were to evolv
Eocene history in b
complicated one, as m
teen million years and
Asia will probably be
Eocene date are alread
faunas. In Africa the
still provide informati
Tertiary. At present it
from the Eocene to th
Again, when we lo
Eocene, the record is
rises from forty-one ir
Of course, it should 1
really due to incomp
Still, the main part of
If the Paleocene wa
as an epoch of consoli
dominant group of la
Table 4. The Eocene (
Epoch
Africa
(faunas
Oligocene
Fayum
Eocene
Paleocene
Mokattam
[PAGE BREAK]
s as large as a
cheek teeth may
sea-cows, which
he proboscideans,
act, found for the
out the same age
y date. The early
the
bient stage
hing of the snout,
eir appearance in
an African origin
been found in the
the lower Eocene
at that early date.
umbers and great
rica, Europe, and
ptation to a com-
ges not projecting
the early whales,
were somewhat
THE EOCENE: EPOCH OF CONSOLIDATION
the snout was elongate and the nostrils were half-way up to the top of the
head. The front teeth were simple cones, but the cheek teeth were power-
ful, shearing structures not unlike those in creodonts; this is especially true
for the middle Eocene archaeocetes and suggests that the whales arose from
some member of the Carnivora or Creodonta which lived on fish, just as
the seals were to evolve much later from swimming carnivores.
Eocene history in both North America and Europe is a very long and
complicated one, as may be seen from Table 4. It spans eighteen or nine-
teen million years and is subdivided into four or five ages. The blanks in
Asia will probably be filled in before long; faunas of early and middle
Eocene date are already known, though not so well as the great late Eocene
faunas. In Africa the situation is less promising, but diligent search may
still provide information on the land faunas of this continent in the early
Tertiary. At present it comes into focus with the Fayum and the transition
from the Eocene to the Oligocene.
Again, when we look at the familial diversity of the mammals in the
Eocene, the record is one of great increase. The number of families known
rises from forty-one in the late Paleocene to ninety-one in the late Eocene.
Of course, it should be remembered that part of the seeming increase is
really due to incompleteness of the Paleocene and early Eocene record.
Still, the main part of the increase is probably real.
If the Paleocene was an epoch of conquest, the Eocene may be regarded
as an epoch of consolidation. The mammals had definitely emerged as the
dominant group of land animals and were now crowning their victory by
Table 4. The Eocene of the World Continent
Epoch
Africa
(faunas)
Asia
North
(faunas)
America
(ages)
Europe
(ages)
Date
(Million
years)
Oligocene
(Mongolia)
Chadronian
Sannoisian
37
Fayum
Irdin Manha Duchesnean
Pondaung
Ludian
hd must have re-
very closely. (A
ury combined the
dred metres long.)
ive than the com-
ratively small and
e large head of a
nally visible neck.
ain creodonts, but
40
Mokattam
Eocene
Uintan
Bartonian
45
Bridgerian
Lutetian
49
Ulan Bulak
Ypresian
Wasatchian
Sparnacian
55
Paleocene
Gashato
Clarkforkian
F
81
[PAGE BREAK]
CF.M.N.H.
ZEUGLODON-A FOSSIL WHALE
[PAGE BREAK]
PALEONTOLOGY
1994
Mesonychid (55 million years ago)
Back to
the Sea
Ambulocetus (50 million years ago)
[PAGE BREAK]
one
IF EVOLUTION IS A MOVIE, it's the job of paleontol
ogists to look for the lost footage. This past year they
came out of the vaults with an awesome director's cut of
of the strangest films ever made: Whale Is Born.
For decades researchers have claimed that whales are
descended from an extinct hyenalike land mammal,
called a mesonychid, that walked back into the sea be
tween 50 and 60 million years ago. (Mesonychids and
all other land mammals are themselves descended from
a fish that crawled out of the sea much earlier.) By -40).
million years ago the transition from four-legged land
animal to fishlike ocean dweller was almost complete.
The evidence is a fossil whale called Prosenglodon. It
was perfectly adapted for life at sea but as University
of Michigan paleontologist Philip Gingerich discov
ered in 1989, it still carried, near the end of its 15-foot
body, a pair of vestigial 6-inch legs.
Rodhocetus (46 million years ago)
Prozeuglodon (40 million years ago)




