
Box 4
Folder 47. Cryptid Whales Beaked Whale Meso Dlodon SP
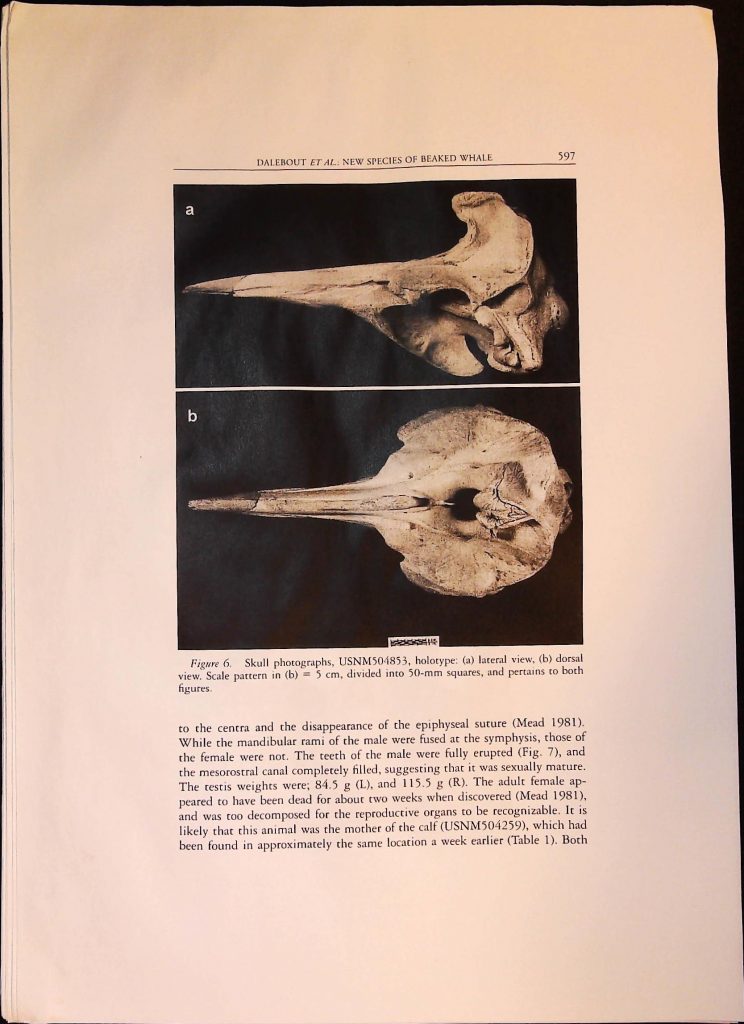
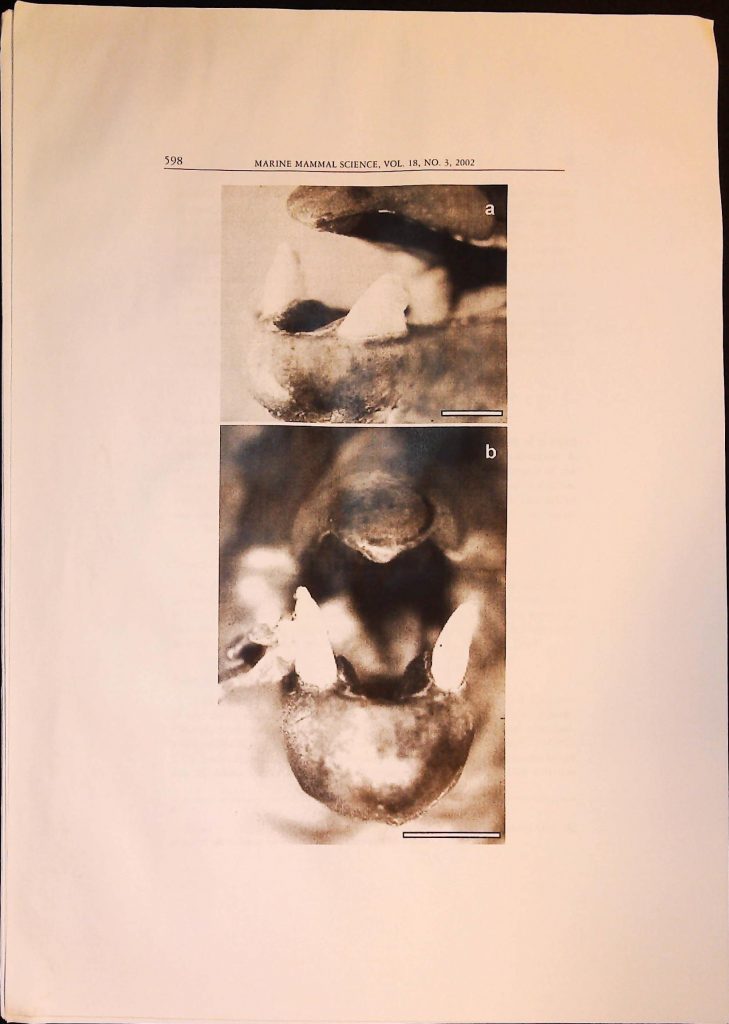
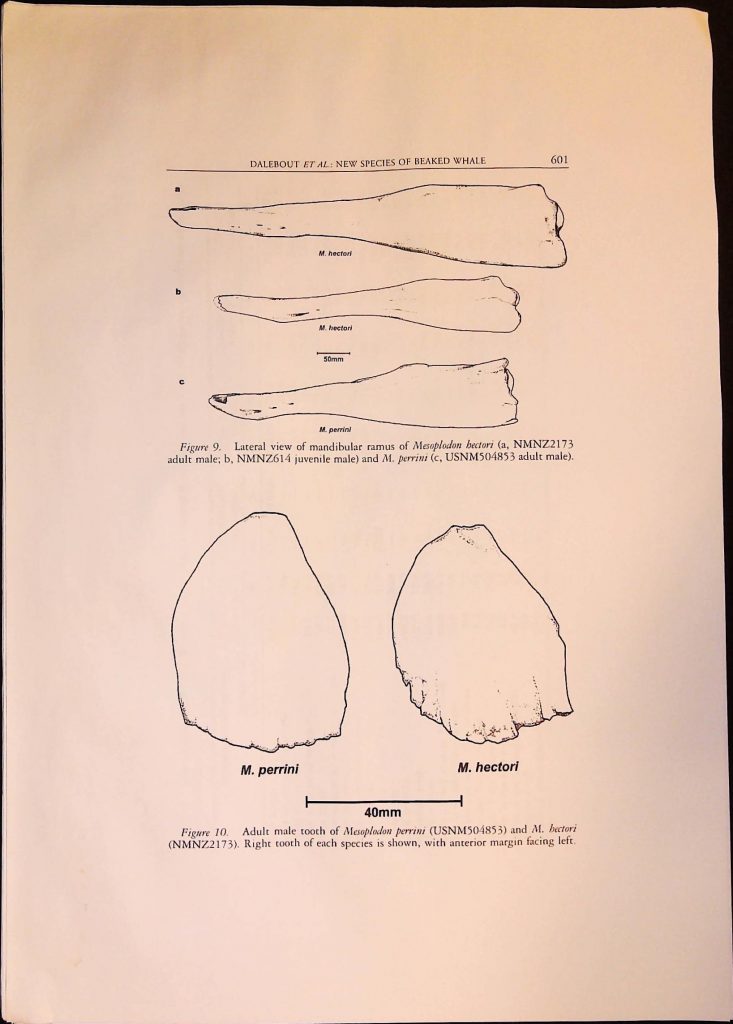
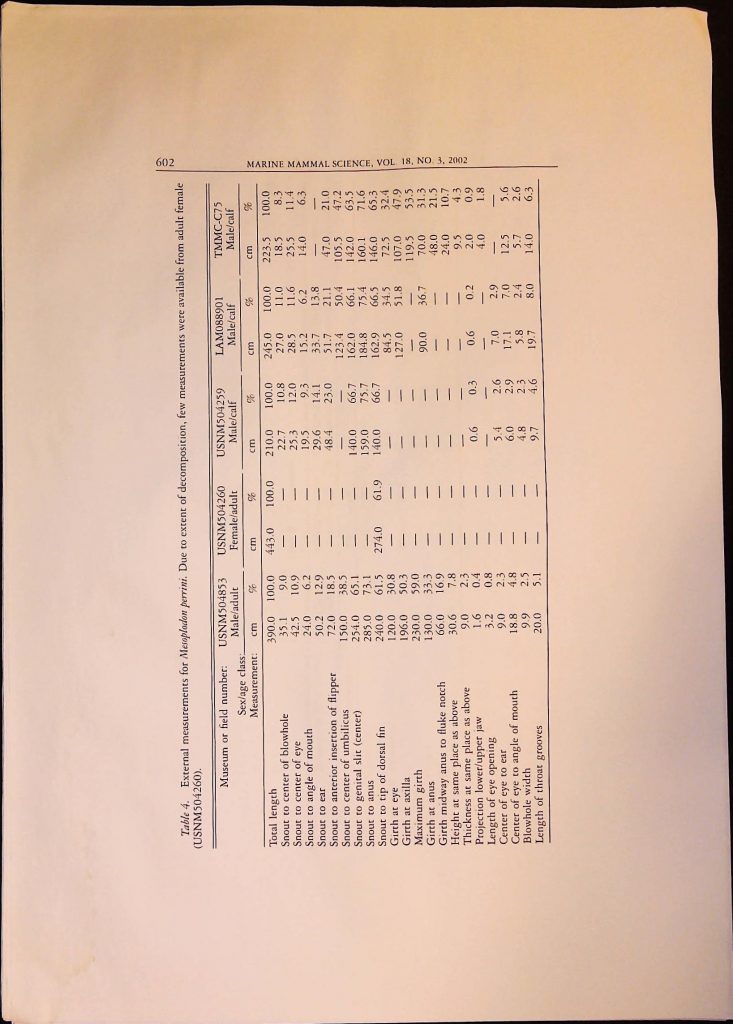
Item 1. Academic Articles







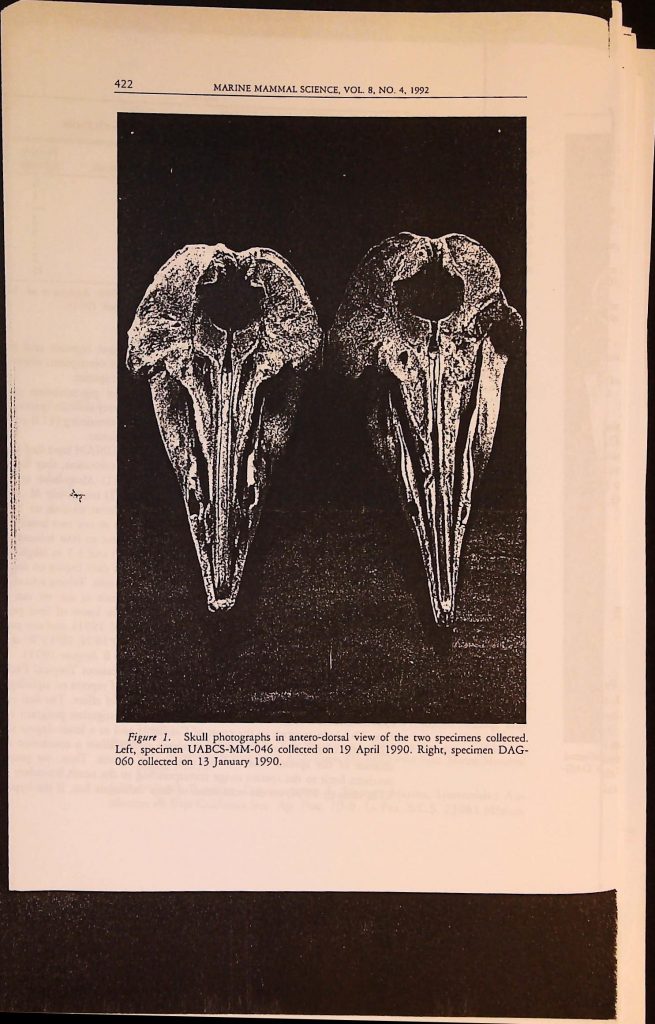



![]()

![]()




![]()













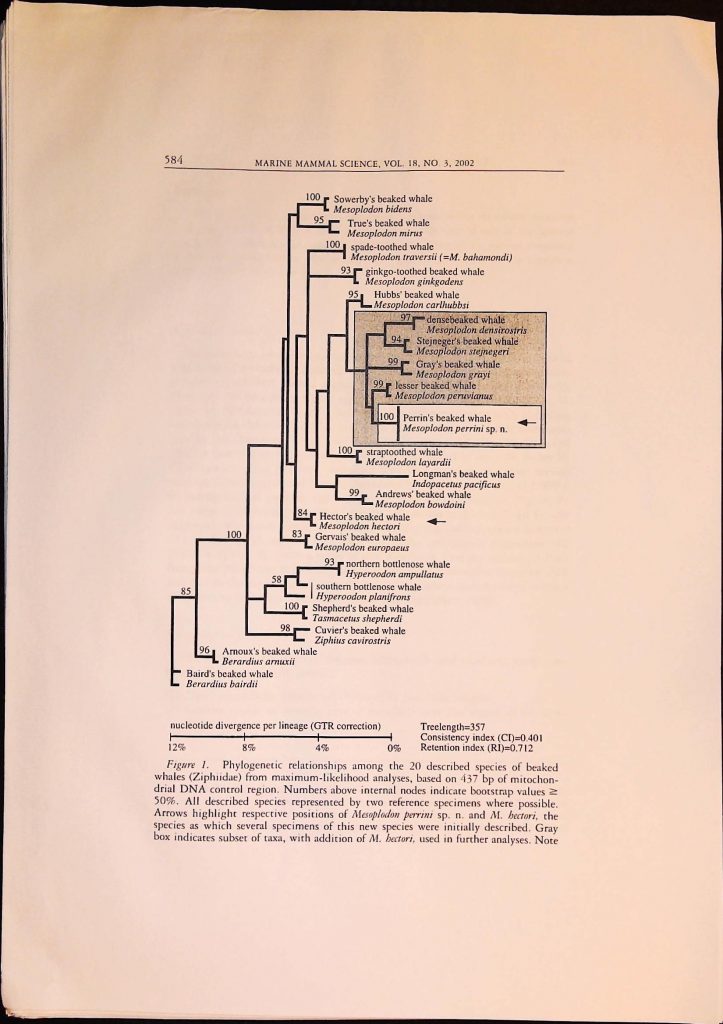
![]()
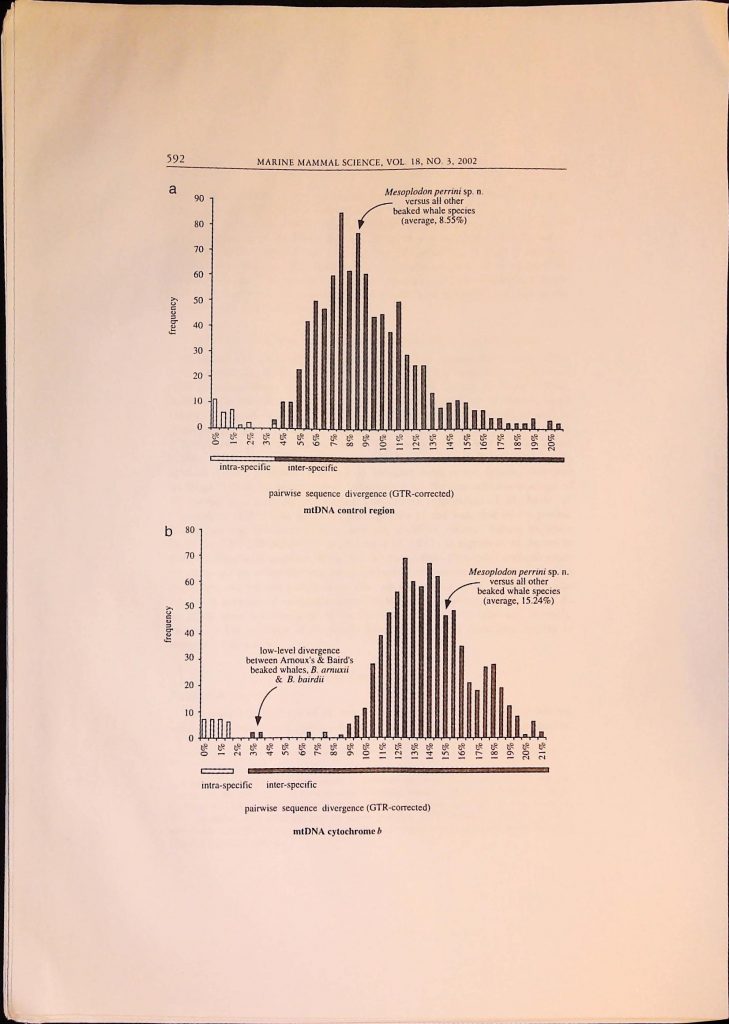
![]()

![]()




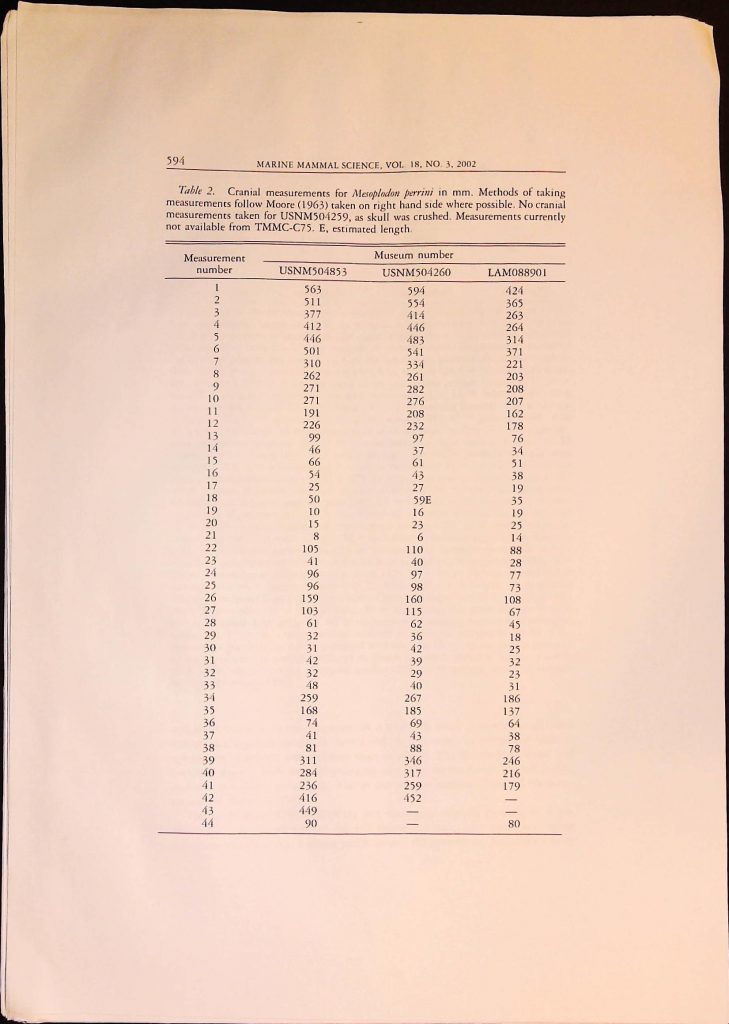

![]()







![]()






Transcribed Text (OCR)
GARY MANGIACOPA ARCHIVE ============================================================ Title: B4F47I1 Slug: b4f47i1 Categories: Cryptozoology, Misplaced Animals Source: https://garymangiacopraarchive.com/b4f47i1 Pages: 63 scanned, 63 extracted OCR: Google Vision API (document_text_detection) Processed: 2026-06-06 ============================================================ 180 A. B. VAN DEINSE AND G. C. A. JUNGE richtius robustus 1). It is of interest to note that Gray in his Catalogue (1866) enumerates Balaena gibbosa as well as Eschrichtius robustus. It was impossible for him to know at that time that they could be identical, for the only fact which was known of Dudley's scrag whale was the external form. The third phase starts with Cope's description of the California gray whale. In 1868 (p. 159) Cope mentioned "that he had the opportunity of examining a portion of the scrag whale of Dudley, Balaena gibbosa of Erxleben, and ascertained that it represented a genus not previously known". The genus was called Agaphelus. He continues: "A second species of the genus was to be found in the "gray whale" of the coasts of California". This species was named Agaphelus glaucus. In the same year (pp. 221-227) he gives a more detailed account of these two species. He stated now that the specimen he called Agaphelus gibbosus was cast ashore on Long Beach, New Jersey, in the autumn of 1866 (two years previously), and that the most important portions for its identification were lost. From the description of the bones, especially the mandibular ramus, the lumbar and caudal vertebrae, which according to Cope resembled those of Balaenoptera rostrata more than those of any other whale and from the description of the colour of the pectoral fins, which were white at the base and black at the end, it seems that Cope was misled and that this whale was a Balaenoptera acuto-rostrala. In 1884 (p. 1124) Cope came also to this conclusion saying: "The Agaphelus gibbosus must be withdrawn from the list of authentic species. The bones which I referred to it are probably those of Balaenoptera rostrata. The characters of the animal in the flesh were given me by persons whom I supposed to be trustworthy, but who may have been mistaken". Cope who had not seen the stranded whale himself was informed of the external form by some people, who stated that the dorsal fin was lacking and that the whale had no plicae on the gular and thoracic regions. Because the animal must have been dead already some time before stranding, we cannot attach too much value to these statements, but it is interesting that Cope immediately combined Dudley's description of the scrag whale with the Pacific whale, which he called Agaphelus glaucus and about whose external form he was informed by Dall. 1) Allen (1916, p. 176) placed Balaenoptera robusta Lilljeborg into the synonymy of Balaenoptera physalus (L.) saying (l.c., p. 179): "Fossil remains, now considered indistinguishable from B. physalus, have been found in Sweden, and formed in 1860 the basis of Lilljeborg's Balaenoptera robusta." It will be clear after our discussion of these bones that this cannot be upheld, the differences between Lilljeborg's specimen and Balaenoptera physalus are too striking. THE CALIFORNIA GRAY WIIALE IN THE ATLANTIC 181 In 1884 Cope thought that the specimen he originally described as Agaphelus gibbosus in reality was a Balaenoptera acuto-rostrata, but before that time in 1869 (p. 15) he had established already a new genus, Rhachia- nectes, for the California gray whale especially on account of the different form of the scapula between his Agaphelus gibbosus (= Balaenoptera acuto-rostrata) and Agaphelus glaucus. The form of the scapula in the first was identical with Balaenoptera, in the latter more like that of Balacna. On the previous page (p. 14) he says in a synopsis that the difference should be the absence of an acromion in Rhachianectes. It is not clear how he came to this statement, for the California gray whale possesses a well developed acromion. When we bear in mind that Cope compared the bones of a California gray whale and a Balaenoptera acuto-rostrata, it will be clear that Agaphelus glaucus remains as the only representative of the genus Agaphelus and that Rhachianectes is merely a synonym. It is remark- able that Cope overlooked this consequence in 1884 and that the name Rhachianectes instead of Agaphelus could hold till now. We believe that the recent finds of the skull-fragments and bones in the Netherlands, which parts seem not to be different in any essential point from those of "Rhachianectes glaucus" make it possible to combine the three phases and to state that Dudley's scrag whale (1725), Balaena gibbosa Erxleben (1777), is the same as Balaenoptera robusta Lilljeborg (1861), Eschrichtius robustus (Lilljeborg) (1865), Agaphelus glaucus Cope (1868) and Rhachianectes glaucus Cope (1869). After this it is clear that the name of this whale must be changed into Eschrichtius gibbosus (Erxleben). We must emphasize that with the bones we have now at hand we cannot give any argument that the Pacific and Atlantic representatives of Eschrichtius should be specifically different. The fact that one species occurs in the northern Pacific as well as in the northern Atlantic is also mentioned for Crustacea, Mollusca, Echino- dermata and Pisces (Berg, 1934, PP. 391-409 and Schmidt, 1928, [AD] pp. 1019-1020). The question whether the populations of the California gray whale in the Atlantic and Pacific Ocean were perhaps connected along the American north coast or (and) the European-Siberian coast cannot be answered yet. Material from these coasts is totally lacking till now. The skull-fragments [AD] found during the Vega expedition (1878-1880) are found too near the Bering Strait to be of any importance for this question. [PAGE BREAK] 178 A. B. VAN DEINSE AND G. C. A. JUNGE Temminckia, 2, 1937 Scrag whale NOMENCLATURE AND DISCUSSION OF LITERATURE The skull-fragments and other bones all point to the fact that the whale now commonly called Rhachianectes glaucus has not always been restricted to the Pacific, but has occurred in the Atlantic too. This at first sight astonishing find made necessary an ample study of the literature and we will show that it is possible now to give an explanation to some facts which were imcomprehensible till now. First of all we believe that this find is the clue to understand what Dudley (1725) meant with the mysterious scrag whale. About the whales described by Dudley, True wrote in his splendid work on the whalebone whales of the Atlantic (1904, p. 37): "All of these are recognizable and have been assigned to their proper places generically, except the scrag whale, which is, and always has been, a stumbling block to cetology." Dudley's description of the scrag whale is the following (1725, p. 258): "The Scrag whale is near a kin to the Fin-back, but, instead of a Fin upon his Back, the Ridge of the Afterpart of his Back is scragged with half a Dozen Knobs or Nuckles; he is nearest the Right Whale in Figure and for Quantity of Oil, his Bone is white, but won't split." The absence of a dorsal fin and the crenulations on the dorsal ridge of the peduncle fully agrees with what is known of Rhachianectes glaucus. Andrews (1914, p. 258) mentions 6-14 crenulations on the peduncle in this species. Also the remark that the bone (whalebone) is white and "won't split" points to this species. Andrews says that the colour of the laminae in this species is yellowish white or yellow and Scammon describes it as "light brown or nearly white" (Andrews, 1914, p. 255). About the structure of the plates Andrews remarks that they are very thick and heavy. We believe, that, now it is known that Rhachianectes also inhabited the Atlantic, Dudley's diagnosis is understandable without difficulties. The scrag whale was said to occur near New England 1). The scrag whale, Knotenfisch (German), Knobbelvisch (Dutch) is found in nearly all older literature on the whales of the Atlantic, but nearly all authors derived their knowledge at second hand and compiled Dudley's description: Klein (1741), Anderson (1746), Brisson (1756), Cranz (1765), Boddaert (1772), Müller (1773). Linneaus did not mention this whale in the 10th and 12th edition of his Systema Naturae. 1) According to Allen (1016, p. 116) the fishermen of the New England coast apply to-day the name scrag whale to small specimens of Eubalaena glacialis (Bonn.). Judging from Dudley's description, however, it is certain that he did not design this species. baleine "Recent and older finds of THE CALIFORNIA GRAY WHALE IN THE ATLANTIC 179 Erxleben in 1777 (p. 610) gave it a Latin name for the first time, grite Balaena gibbosa, with diagnosis: "Figura Mysticeti, at gibbi in dorso sex, Alautique N., laminaeque corneae albae. Species subobscura". He quoted most of the 18 ficcle, carlier authors in the synonymy. Much news was not added to the know- ledge of this whale by Erxleben either and it seems that he too derived his knowledge from others. Gmelin mentioned in 1789 (p. 225) Balacna gibbosa. Under this name the scrag whale and the humpback (Megaptera) are combined under a and b. His references also are of second hand. It seems of less importance to enumerate all the later authors, who mentioned Balaena gibbosa in their papers, the most important are Lace- pède (1804), Desmarest (1822), G. Cuvier (1823), F. Cuvier (1836), Brandt and Ratzeburg (1829), Gray (1846, 1850). Most of them do not give much news and many of them combined the scrag whale and the humpback under Balacna gibbosa. F. Cuvier thought it possible that Dud- ley's scrag whale was a rorqual, which had been mutilated, Dudley's note he considered to be insignificant (1836, p. 355). This view was criticized already by Gray (1846, p. 16; 1850, p. 19), who thought the scrag whale to be "a Balaena, probably well known formerly". He also criticized the lumping together of the scrag whale and the humpbacked whale in one genus as was done by many authors and said that they did not understand Dudley's description of the belly of the latter, "being reeved', that is, plaited". He established the genus Megaptera for the humpbacked whales (1846, p. 16). Though much was written about the scrag whale till then, hardly any- thing was known about this species. A second phase in the history of this whale starts independently from the former in 1860. In this year Lilljeborg read a paper in Copenhagen about the find of a subfossil whale near Gräsö in Sweden and he named it Balaenoptera robusta. In 1862 he gave a detailed survey in Swedish and repeated it in English in 1867 with good plates of many of the bones. In 1864 Gray (p. 350) referred this whale to the genus Megaptera and established the subgenus Eschrichtius for it. In 1865 he received a cervical vertebra from the Babbacombe bay, England, and Lilljeborg, who compared it with the cervical vertebrae of his Balaenoptera robusta, considered them identical. Gray (1865, p. 42) raised Eschrichtius to generic rank now and thought this genus was related with Megaptera. He mentions as especially characteristic for this genus the very low processus coronoideus of the lower jaw, the small bodies of the cervical vertebrae, which are suborbicular-quadrangular and possess a wide spinal canal. In his paper of 1867 Lilljeborg too called this whale Esch- [PAGE BREAK] Statut Inconnu BALEINES À BEC 113 Population Inconnue Menaces Inconnues IDENTIFICATION .coloration foncée et claire bande blanc crème bec assez court ⚫ tête relativement plate • melon petit mais bien visible dimorphisme sexuel très marqué ⚫petits groupes en formation serrée ⚫léthargique et lent ⚫ comportement aérien réduit probablement pas d'échancrure médiane nageoire dorsale basse, triangulaire (variable) bord de fuite rectiligne ou légèrement falciforme base large ⚫ corps couvert de cicatrices, surtout derrière le bord de la bande NAGEOIRE CAUDALE licrée brun-noir ou chocolat tronçon caudal ⚫ assez étroit Nouveau-ne Inconnu Multe Em 55m MÅLE (RECONSTITUTION) EAUX PROFONDES DU PACIFIQUE ORIENTAL TROPICAL EN GENERAL AU LARGE Poids à la naissance Poids adulte Inconnu Inconnu AIRE DE RÉPARTITION La carte indique la zone où l'animal a été le plus souvent signalé, mais les observations sont si peu nombreuses qu'elles ne reflètent pas nécessairement la distribution réelle de l'espèce, qui est l'une des plus fréquemment signalees, concernant le genre Mesoplodon, dans les eaux tropicales du grand large du Pacifique oriental. Elle a été vue à ce jour au large des côtes du Guatemala, du Salvador, du Mexique, du Costa Rica, du Nicaragua, de Panama, de Colombie, de l'Equateur et du Pérou, essentiellement dans les eaux profondes, toujours dans des zones assez chaudes, d'environ 27 'C. On ne sait pas quel est son statut, mais le nombre des animaux signalés indique qu'elle n'est pas particulièrement rare pour une baleine à bec. Régime alimentaire [PAGE BREAK] L CEI LONATURE BALEINES DAUPHINS ET MARSOUINS MARK CARWARDINE Illustrations de MARTIN CAMM Conseiller DR PETER EVANS (Sea Watch Foundation, Université d'Oxford, R.U.) MASON WEINRICH (Cetacean Research Unit, Massachussets, E.U) BORDAS 1996 [PAGE BREAK] GOSSE (Philip Henry) The Romance of Natural History CLondon, Nisbet, 1860) 366 THE GREAT UNKNOWN. rally from fifty to three hundred fathoms' depth of water within a boat's length of the shore. How could a carcase or a skeleton be cast up here, even if it floated? But, secondly, as to facts. Is it true, that of all the larger oceanic animals we find the carcases or skeletons cast up on the shore? Is it true even of the Cetacea, whose blubber-covered bodies invariably ensure their floating, and whose bones are so saturated with oil that they are but little heavier than water? In September, 1825, a cetacean was stranded on the French coast which was previously unknown to natural- ists. It was so fortunate as to fall under the examina- tion of so eminent an zoologist as De Blainville; and hence its anatomy was well investigated. It has become cele- brated as the Toothless Whale of Havre (Aodon Dalei). Yet no other example of this species is on record; and, but for this accident, a whale inhabiting the British Channel would be quite unrecognised. Of another whale (Diodon Sowerbyi), likewise British, our entire knowledge rests on a single individual which was cast on shore on the Elgin coast, and was seen and described by the naturalist Sowerby. There is a species of sperm-whale (Physeter tursio) affirmed to be frequently seen about the Shetland Islands; a vast creature of sixty feet in length, and readily dis- tinguishable from all other Cetacea by its lofty dorsal, and, according to old Sibbald, by other remarkable pe- culiarities in its anatomy. Yet no specimen of this huge creature has fallen under modern scientific observation; GOSSE'S DELPHINORHYNCHUS. 367 and zoologists are not yet agreed among themselves, whether the High-finned Cachalot is a myth or a reality! M. Rafinesque Smaltz, a Sicilian naturalist, described a Cetacean which, he said, he had seen in the Mediterranean, possessing two dorsals. The character was so abnormal that his statement was not received; but the eminent zoologists attached to one of the French exploring expedi- tions, MM. Quoy and Gaimard,--saw a school of cetacea around their ship in the South Pacific, having this extra- ordinary character,-the supernumerary fin being placed on the back of the head. Here is the evidence of com- petent naturalists to the existence of a most remarkable whale, no carcase of which, no skeleton, has ever been recognised. The last example I shall adduce is from my own ex- perience. During my voyage to Jamaica, when in lat. 19° N., and long. from 46° to 48° W., the ship was sur- rounded for seventeen continuous hours with a troop of whales, of a species which is certainly undescribed I had ample opportunity for examination, and found that it was a Delphinorhynchus, thirty feet in length, black above and white beneath, with the swimming paws white on the upper surface, and isolated by the surrounding black of the upper parts,-a very remarkable character. This could not have been the Toothless Whale of Havre; and there is no other with which it can be confounded. Here, then, is a whale of large size, occurring in greut numbers in the North Atlantic, which on no other occa- sion has fallen under scientific observation. [PAGE BREAK] 420 MARINE MAMMAL SCIENCE, VOL. 8, NO. 4, 1992 LITERATURE CITED BOYD, I. L., AND C. D. Duck. 1991. Mass changes and metabolism in territorial male Antarctic fur seals (Arctocephalus gazella). Physiological Zoology 64:375-392. COSTA, D. P., B. J. LE BOEUF, A. C. HUNTLEY AND C. L. ORTIZ. 1986. The energetics of lactation in the northern elephant seal, Mirounga angustirostris. Journal of Zoology, London 209:21-33. JOHNSON, R. R., AND D. J. FARRELL. 1988. The prediction of body composition in poultry by estimation in vivo of total body water with tritiated water and deuterium oxide. British Journal of Nutrition 59:109-124. ØEN, E. O. 1982. A new darting gun for the capture of wild animals. Nordic Veterinary Medicine 34:39-43. ØEN, E. O. 1990. Trials of chemical immobilization of minke whales with etorphine hydrochloride 1989. Reports of the International Whaling Commission (SC/42/ NHMi18). 891. REILLY, J. J., AND M. A. FEDAK. 1990. Measurement of the body composition of living grey seals by hydrogen isotope dilution. Journal of Applied Physiology 69:885- RUMPLER, W. V., M. E. ALLEN, D. E. ULLREY, R. D. EARLE, S. M. SCHMITT AND T. M. COOLEY. 1987. Body composition of white-tailed deer estimated by deuterium oxide dilution. Canadian Journal of Zoology 65:204–208. VILJOEN, J., S. E. COETZEE AND H. H. MEISSNER. 1988. The in vivo prediction of body composition in Boer goat does by means of tritiated water space technique. South African Journal of Animal Science 18:63-67. CHRISTIAN LYDERSEN, University of Oslo, Department of Biology, Division of General Physiology, P.O. Box 1051 Blindern, N-0316 Oslo 3, Norway; DAVID GRIFFITHS, Norwegian College of Veterinary Medicine, Department of Anatomy, [AD] P.O. Box 8146 dep., N-0033 Oslo 1, Norway; and IAN GJERTZ AND ØYSTEIN WIG, Norwegian Polar Research Institute, P.O. Box 158, N-1330 Oslo Luf- thavn, Norway. Received October 15, 1991. Accepted January 12, 1992. MARINE MAMMAL SCIENCE, 8(4):420-425 (October 1992) 1992 by the Society for Marine Mammalogy FIRST RECORD OF THE PYGMY BEAKED WHALE MESOPLODON PERUVIANUS IN THE NORTH PACIFIC Two specimens of the genus Mesoplodon were found in the Bahía de La Paz, Baja California Sur, México. The first one was found on 13 January 1990, on a beach to the side of the road from La Paz City to San Juan de la Costa (24°24'N, 110°38'W). The carcass was rotten, the length was estimated at 3.4 m, and the skull was collected and deposited in Aurioles' personal collection in La Paz, B.C.S. (DAG 160). On 19 April 1990, an almost complete skeleton of a second specimen was collected from a sandy beach on the west coast of the Isla Espiritu Santo (24°25'N, 110°25'W). The length of this animal was cal- Strictement réservé à l'usage privó (Loi n° 57293 du 11.3.57, art. 41,0) [PAGE BREAK] 421 ial male 392. ergetics Coology, ition in uterium terinary orphine C/42/ f living 9:885- T. M. iterium of body South ion of DAVID tomy, YSTEIN 2. E Luf- Paz, 0, on Costa t 3.4 on in leton of the Scal- NOTES culated as 3.3 m. The skeleton was deposited in the Museo de Zoología of the Universidad Autónoma de Baja California Sur, La Paz, B.C.S. (UABCS- MM-053). Because of the distal fusion of the premaxilla and maxilla, the degree of filling of the mesorostral canal, and the fusion of the vertebral epiphysis to the centra of the vertebrae, it was concluded that both animals were physically mature. Based on the length of the animals and their particular skull characteristics such as tooth size and shape, tooth position, and synvertex configuration (Fig. 1), among others, they were initially thought to be a new species, and later determined to be Mesoplodon peruvianus, the species described recently by Reyes et al. (1991). These records are the first of M. peruvianus for the North Pacific, and, in fact, the first outside of the Peruvian coasts, where there is information on ten specimens, including the holotype, which have either stranded or been captured between 11°12'S and 15°19'S from 1976 to 1989 (Reyes et al. 1991). The only other mesoplodont species recorded for the Mexican Pacific is M. ginkgodens from a skull found on the Playa Mal Arrimo near the mouth of the Laguna Ojo de Liebre, Baja California Sur, in December 1980 (Leatherwood et al. 1982). Besides M. peruvianus, 6 of the 13 Mesoplodon species are present in the North Pacific Ocean (Leatherwood et al. 1982, Mead 1989), with the possibility of one more undescribed species (Pitman et al. 1987, 1988), for the Eastern Tropical Pacific (ETP) (Table 1). yr There was almost no information about the ziphiids from the ETP until the last 15 when important data were obtained by trained observers during marine mammal survey cruises for the Southwest Fisheries Center, and by observers on tuna purse seiners during their fishing activities (Perrin et al. 1985, Pitman et al. 1988). Information on ziphiids has also been obtained from the long term Marine Mammal Research Program of the Universidad Nacional Autónoma de México (UNAM), in the Bahía de Banderas, Nayarit on the mainland coast of México since 1981; from the Marine Mammal Research Program of the Uni- versidad Autónoma de Baja California Sur (UABCS), in the Bahía de la Paz, Península de Baja California since 1987; and from the work on stranded marine mammals in the Bahía de La Paz during the last 12 yr by one of us (DAG). As a result of these investigations, there have been 143 positive and 43 probable sightings of mesoplodonts of at least three different species in the ETP (Pitman et al. 1988, in litt. 8 August 1991): (1) Mesoplodon densirostris (4 positive sightings). (2) Mesoplodon sp. "A" (24 positive sightings). This un- identified species was described by Pitman et al. (1987). The animals reach a maximum estimated length of 5.0-5.5 m, larger than the average for Mesoplodon. The species appears to be sexually dimorphic, with a very distinctive color pattern for the presumed adult male, with a broad white swath that originates in the head area and runs postero-ventrally down both sides. The resulting chevron pattern contrasts sharply with the otherwise black body. (3) Mesoplodon grayi (2 positive sightings). This species has an extremely long, narrow rostrum. The [PAGE BREAK] Jorg 422 MARINE MAMMAL SCIENCE, VOL. 8, NO. 4, 1992 Figure 1. Skull photographs in antero-dorsal view of the two specimens collected. Left, specimen UABCS-MM-046 collected on 19 April 1990. Right, specimen DAG- 060 collected on 13 January 1990. [PAGE BREAK] :0 NOTES 45 32 Table 1. Records of Mesoplodon spp. in the North Pacific. Species M. stejnegeri M. carlhubbsi Stranded animals Captured animals Positive sightings M. ginkgodens M. hectori M. densirostris M. peruvianus M. grayi M. sp. 25 "A" 423 9.9 ut of pio [D YST 12. [1] Pa 0, d Cost letofollected. t 3. on i of th s cal 2 DAG- Sources: Leatherwood et al. 1982; Pitman et al. 1987, 1988; Aguayo et al. 1988; Mead et al. 1988; Mead 1989; R. L. Pitman (in litt. 8 August 1991). lower jaw contrasts markedly with the dark gray upper rostrum and has a distinctive pale patch just behind the eye. Based on the descriptions above, we can assume M. peruvianus is different from these three species. Owing to the small size and not very distinctive pigmentation of M. peruvianus, we can expect the field identification to be very difficult (particularly prior to the species description), therefore several of the remaining (113) positive sightings of Mesoplodon sp. could have been M. peruvianus. In Bahía de Banderas, México, researchers from the UNAM have had several encounters with mesoplodonts and, based on size and coloration, they found at least three different species (Aguayo et al. 1988): (1) Mesoplodon sp. "A" described before, (2) tentatively M. ginkgodens, and (3) tentatively M. hectori. One of us (JUR) had the opportunity to observe the latter animals on 13 July 1986. Two individuals came to the surface very close to our two small boats, and we could aproach them to within 4-6 m for three to four minutes. The blow was inconspicuous, their length was between 3.0 and 3.5 m (slightly less than half the length of the boats), their coloration was dark brown on the back and sides and lighter below, and no teeth or scars were seen. Taking into account the difficulty of positive identification of these animals at sea we can now, tentatively, consider them as M. peruvianus. Thus we know of four possible sightings of this species in the South Pacific (Reyes et al. 1991), and one positive sighting in the North Pacific off southern Mexico at 10°58'N, 98°15'W of three animals, two adults and a calf (R. L. Pitman, in litt. 8 August 1991). Based on the above information we propose the Eastern Tropical Pacific as the distribution area for M. peruvianus; the paucity of reports on sightings and strandings in this area can be attributed to the lack of effort. The first reports appeared after initiation of an active stranding investigation program in this area. This is in contrast to the coast of California and to a lesser degree to Baja California coasts, where the extensive survey efforts allow a conclusion of the absence of the species based on an absence of reports. Thus, we propose a northern limit to the species range corresponding to the north boundary of the ETP, close to 25°N, on the west coast of Baja California Sur. If the hypothesis [PAGE BREAK] 424 MARINE MAMMAL SCIENCE, VOL. 8, NO. 4, 1992 of the distribution in the ETP is correct, the mesoplodonts found on the Peruvian coast should be close to the limit of their southern range (15°S). This distribution is similar to the one proposed for Mesoplodon sp. "A" (Pitman et al. 1987, 1988; Mead et al. 1988), and the distribution of the tropical dolphins (Stenella spp. and Delphinus) in the ETP (Perrin et al. 1985). Based on the geographical distribution of the M. peruvianus records, and their probable foraging in mid and deep waters, it seems they have a distribution similar to the striped and the common dolphins, which occur in equatorial and subtropical waters with relatively large seasonal changes in the surface temperature, depth of the thermocline, and upwelling. ACKNOWLEDGMENTS We thank the staff of the Marine Mammal Program of the School for Field Studies, especially Dr. Gary Miller for finding information on the UABCS-MM-053 specimen. Dr. D. Wartzok, the editor, reviewed the manuscript and corrected the English. Dr. J. suggestions. Mead, Dr. R. Brownell and R. Pitman critically reviewed the manuscript and made LITERATURE CITED AGUAYO L., A., D. AURIOLES G., J. URBAN R., M. SALINAS Z., O. VIDAL AND L. T. FINDLEY, 1988. Beaked whales in Mexican waters. Paper SC/40/SM/13 presented to the Scientific Committee of the IWC Annual Meeting, May 1988. LEATHER WOOD, S., R. R. REEVES, W. F. PERRIN AND W. E. EVANS. 1982. Whales, dolphins and porpoises of the eastern North Pacific and adjacent waters: a guide to their identification. NOAA Technical Report, NMFS Circular 444. 245 pp. MEAD, J. G., J. E. HEYNING AND R. L. BROWNELL, JR. 1988. Distribution and ex- ploitation of beaked whales in the Northern Hemisphere. Paper SC/40/SM/21 presented to the Scientific Committee of the IWC Annual Meeting, May 1988. MEAD, J. G. 1989. Beaked whales of the genus Mesoplodon. Pages 349-430 in S. H. Academic Press, N.Y. Ridgway and R. J. Harrison, eds. Handbook of marine mammals, volume 4. PERRIN, W. F., M. D. SCOTT, G. J. WALKER AND V. L. CASS. 1985. Review of geographical stocks of tropical dolphins (Stenella spp. and Delphinus delphis) in the NMFS 28. 28 pp. eastern North Pacific. U.S. Department of Commerce, NOAA Technical Report PITMAN, R. L., A. AGUAYO L. AND J. URBAN R. 1987. Observations of an unidentified 3:345-352. beaked whale (Mesoplodon sp.) in the eastern tropical Pacific. Marine Mammal Science PITMAN, R. L., D. W. K. Au, M. SCOTT AND J. M. COTTON. 1988. Observations of beaked whales (Ziphiidae) from the eastern tropical Pacific Ocean. Paper SC/40/ 1988. SM/14 presented to the Scientific Committee of the IWC Annual Meeting, May REYES, J. C., J. G. MEAD AND K. VAN WEAREBEEK, 1991. A new species of beaked Science 7:1-24. whale Mesoplodon peruvianus sp. n. (Cetacea: Zhipiidae) from Peru. Marine Mammal JORGE URBÁN-RAMÍREZ, Departamento de Biología Marina, Universidad Au- tónoma de Baja California Sur. Ap. Post. 19-B, La Paz, B.C.S. 23081 México; [PAGE BREAK] PERSPECTIVES SCIENTIFIQUES Un cousin des dauphins Le mésoplodon nain est la treizième espèce du genre. In 1976, le zoologue James Mead de la Smithonian Institution, découvre les restes d'un cétacé inconnu sur un marché au poisson du Pérou. Un travail de longue haleine sera nécessaire jusqu'à ce qu'une nouvelle espèce soit officielle- ment enregistrée sous le nom de méso- plodon nain (Mesoplodon peruvianus Reyes, Mead & van Waerebeek, 1991). James Mead et ses collègues du Cen- tre péruvien d'étude des cétacés mettent près de 15 ans pour collecter un nombre suffisant de spécimens, pris dans des filets de pêche ou échoués sur des plages: l'animal semble rare et les zones où il vit sont peu fréquentées par les cétologues. Une dizaine de spécimens seulement a été récupérée, et il a fallu attendre 1985 pour en obtenir un complet. Le Musée océanographique de La Rochelle présente actuellement une exposition autour du crâne de l'un de ces animaux qu'il a acquis pour enrichir ses collections. Ce spécimen a été capturé en janvier 1992 au large de la localité de Pucusana au Pérou. Les mésoplodons sont des odonto- cètes, des cétacés à dents, membres de la famille des ziphiidés. Les odontocètes les plus connus sont les dauphins (famille des delphinidés), les cachalots (famille des physétéridés), et les marsouins (fa- mille des phocénidés). La première espèce de mésoplodon fut répertoriée en 1804. Il s'agit du «mésoplodon de Sowerby» qui vit principalement en Mer du Nord. Les autres ont été découvertes tout au long du XIXe et du XXe siècle. Les deux dernières l'avaient été en 1958 et 1963. Le mésoplodon nain porte à treize le nombre d'espèces du genre, une sorte de record: les autres genres de la famille des ziphiidés ne comptent qu'une ou deux espèces chacun. Les mésoplodons ne possèdent qu'une paire de dents, implantée sur la mâchoire inférieure. Leur aileron dorsal est situé très en arrière, et leur queue ne porte pas d'encoche centrale. Le méso- plodon nain est plus petit que les autres espèces. Le plus grand spécimen est un mâle de 3,72 mètres de long alors que les autres mésoplodons ont des tailles comprises entre 4 et 6 mètres. La posi- tion de ses dents et d'autres caractères crâniens le différencient également. Son dos est de couleur gris brun: la pigmen- tation s'éclaircit sur les flancs pour deve- nir presque blanche sur le ventre. L'enregistrement d'une nouvelle espèce est plus fréquent pour des petits animaux comme les rongeurs que pour des mammifères de grande taille Le peu d'explorations menées dans le Pacifique tropical sud où vit le mésoplodon nain explique sans doute qu'il n'ait pas été identifié plus tôt. Selon Daniel Robineau, du Muséum national d'histoire naturelle, la découverte d'une nouvelle espèce au sein d'un groupe restreint comme les cétacés, qui en compte environ 80, mon- tre que notre connaissance de cette faune n'est pas exhaustive: «L'image très populaire des dauphins laisse penser au public que l'on connaît tous les céta- cés. Nombre d'espèces actuelles, que l'on trouve dans différentes régions du Globe, présentent des formes locales encore mal connues dont certaines seront sans doute considérées comme des espèces distinctes. Par ailleurs la découverte d'es- pèces entièrement nouvelles n'est pas à exclure il existe encore de nombreuses zones océaniques peu explorées. >> Un mésoplodon marin, capturé accidentellement par des pêcheurs péruviens. La physique dans un plan Élasticité et ruptures d'un Cristal à deux dimensions. L es physiciens aiment à simplifier la nature pour dépister les phénomènes nouveaux. Si les cristaux ont habituelle- ment trois dimensions, l'étude des carac- téristiques à deux dimensions permet d'isoler des phénomènes intéressants, de valider les théories qui s'y rapportent et d'imaginer ensuite des applications techniques. Les cristaux à deux dimensions sont constitués par des molécules confinées à la surface de l'eau. Une partie hydro- phile de ces molécules «amphiphiles>> réside dans l'eau, et une autre partie hydrophobe se maintient hors de l'eau. Nous avons utilisé des molécules d'acide stéarique modifié dont les deux extrémi- tés sont inégalement hydrophobes. Selon la température et la pression (bidi- mensionnelle), les molécules sont dans les états gazeux, liquide ou solide. Dans les phases gazeuse et liquide, les deux groupes hydrophiles sont dans l'eau. Dans la phase solide, la molécule est redressée, et seul le groupe acide est dans l'eau. Ainsi la solidification corres- pond à un changement notable de l'aire du film (d'un facteur presque égal à trois); en comprimant lentement le film liquide, on fait croître des cristaux dans la phase liquide Une propriété optique permet de suivre la progression de la phase solide : celle-ci émet une fluorescence jaune quand elle est éclairée en lumière bleue, tandis que les phases liquide et gazeuse n'émettent presque pas. Lorsque la compression du film est lente, les monocristaux qui croissent dans la phase liquide sont des aiguilles rectangulaires d'une molécule d'épais- seur, de plus d'un millimètre de longueur et de quelques centièmes de millimètre de largeur. Pour mesurer la flexibilité de ces bandes cristallines lors d'une défor- mation dans le plan de l'eau, on utilise un dispositif constitué de deux aiguilles en fibre de verre fixes et d'une aiguille mobile et flexible, qui pousse le cristal bidimen- sionnel en son milieu (voir la figure). On mesure au microscope le fléchissement des aiguilles de verre pour connaître la force appliquée et le fléchissement du cristal sous l'action de cette force. Après l'élimination de nombreux arte- facts expérimentaux dus aux forces capil- POUR LA SCIENCE N°201 JUILLET 1994 25 25 [PAGE BREAK] MARINE MAMMAL SCIENCE, 3(4):345-352 (October 1987) ©1987 by the Society for Marine Mammalogy ra novaeangliae eds. Handbook Academic Press, ey, ed. Marine ch Press, Seat- tober 2, 1986 arch 23, 1987 OBSERVATIONS OF AN UNIDENTIFIED BEAKED WHALE (MESOPLODON SP.) IN THE EASTERN TROPICAL PACIFIC ROBERT L. PITMAN Southwest Fisheries Center, [AD] P.O. Box 271, La Jolla, California 92038 ANELIO AGUAYO L. JORGE URBAN R. Facultad de Ciencias, UNAM, Apartado Postal 70-572, C.P. 04510, México, D.F., México ABSTRACT Multiple sightings of a distinctive but unidentified species of beaked whale have been made in the eastern tropical Pacific. The unidentified whale has two color morphs: a conspicuously marked black and white form (judged to be larger), and a uniformly gray-brown form. Maximum length estimates have been 5-5.5 m. Other features include a relatively flat head, with a small, distinct melon; a moderately long beak; and a low, wide-based, triangular dorsal fin. On most animals the trailing edge of the dorsal fin is only slightly falcate and often appears straight. On the black and white morph, a broad white or cream-colored swathe originates immediately posterior to the dorsal surface of the head and runs posterio-ventrally on either side of the animal. The prevalence of scarring on the black and white animals suggests sexual dimorphism and that these larger, more conspicuously marked animals are adult males, while the smaller, browner, unscarred animals are females and young. Possibilities for identification include: 1) a well-marked race of a known Mesoplodon sp., 2) Mesoplodon (Indopacetus) pacificus or 3) an undescribed species. Key words: Mesoplodon, unidentified beaked whale, eastern tropical Pacific. In this paper we report on multiple sightings of a distinctive but as yet unidentified species of beaked whale from the eastern tropical Pacific (ETP). A morph of this whale that we assume is the adult male (see below) is conspicuously description of a marked and eminently identifiable in the field. Despite this, it does not fit the any known species of ziphiid. We provide here a field description of the whale, identify its range and comment on its identity. 345 [A th O b V [PAGE BREAK] PITMAN ET AL: UNIDENTIFIED BEAKED WHALE 347 [A th I a S Figure 2. b Photographs of the unidentified beaked whale from Bahia de Banderas, Jalisco, Mexico. The animal shown in 2a and 2b, foreground, is presumably an adult male; notice the scratches behind the white area. Photos by P. Hernández and J. L. López. The distinctive dorsal fin is low, wide-based and triangular. On most animals the trailing edge of the fin is only slightly falcate and often appears straight. Most of the dorsal surface of the black and white form, including the dorsal fin, is dark and has been variously described as blackish, black/brown, chocolate brown or dark olive brown. This is similar to the range in color variation noted for short-finned pilot whales (Globicephala macrorhynchus) in the eastern Pacific. Pilot whales in the field are generally described as jet black, but in the ETP they often appear greenish brown. Whether these perceived differences are the result of lighting effects, as seems likely, or some actual physical differences, is not known. For the time being, the unidentified whale is best described as black and white. m b [PAGE BREAK] 346 30 N MARINE MAMMAL SCIENCE, VOL. 3, NO. 4, 1987 Hawallan Islands ⚫ Subtropical 20° Seasonally Tropical 10° Tropical. Equatorial 10° 20°S 160° 150° Subtropical 140° 130° 120° 110° Mexico 100° O Sightings Tentative sightings Survey vessel noon positions (500 10x1) South America 90° 60° 70° Figure 1. Sightings of the unidentified beaked whale in the eastern tropical Pacific relative to surface water types (after Wyrtki 1967). SIGHTINGS Figure 1 shows 24 positive and 8 tentative sightings of the unidentified whale. All except one were made by trained observers during marine mammal survey cruises for the Southwest Fisheries Center in the central and eastern tropical Pacific (see Perrin et al. 1983). Noon positions of survey vessels during these cruises are also plotted in Figure 1 and indicate the extent of survey coverage. The remaining sighting (Fig. 2) was photographed during a marine mammal identification course taught by two of us (AAL and JUR) at Facultad de Ciencias, Universidad Nacional Autonoma de Mexico (UNAM). Figure Jalisco, 1. male; no DESCRIPTION The unidentified whale has two distinct color morphs: a conspicuously marked black and white form and a relatively nondescript morph that appears to be uniformly gray-brown (Figs. 2 and 3). When they have been seen together, the black and white animal has been judged by several observers to be the larger of the two. Estimates of maximum length have most often been between approximately 5-5.5 m, making it an average-to-slightly-large-sized Mesoplodon (see Mead 1984). The head is relatively flat, as in most mesoplodonts, with a small but distinct melon. The beak is moderately long, appearing similar in size to that of M. densirostris. On one occasion an observer reported that the mandible was paler than the upper part of the beak on one of the black and white animals. The the trail Most fin, is d brown for shor Pilot w they of result d not knea and w [PAGE BREAK] PITMAN ET AL.: UNIDENTIFIED BEAKED WHALE 351 forms a unique biotic province in the eastern Pacific (Reid 1962, Wyrtki 1967, McGowan 1972, Au and Perryman 1985). The range of the unidentified whale is also nearly identical to the range of the eastern spinner dolphin (Stenella longirostris), a pantropical species with a well-marked form endemic to the eastern tropical Pacific (Perrin et al. 1985). From the evidence above, it seems likely that a distinct, possibly endemic, species of Mesoplodon also occurs in the eastern tropical Pacific. Possibilities for identification of the unidentified whale include the following: 1) It may be a well-marked race of a known species of Mesoplodon. 2) Mesoplodon (Indopacetus) pacificus is currently the rarest extant species of whale known and has never been identified in the flesh. It is known from only two skulls: one from Queensland, Australia, and one from Somalia, northeast Africa. It is possible, however, that these records represent extralimital strays from a localized population inhabiting a little-explored area, such as the eastern tropical Pacific. 3) It may be an undescribed species. The identity of the whale will remain unknown until a stranded specimen is collected. ACKNOWLEDGMENTS We would like to thank Jim Cotton, Gary Friedrichsen and Scott Sinclair for their capable assistance in the field. David Au, Jim Mead, John Heyning and especially William Perrin improved the manuscript with their comments. We thank P. Hernández and J L. López for use of their photographs, Henry Orr for drafting Figure 1, and we gratefully acknowledge the talents of Lou Silva in preparing Figure 3. LITERATURE CITED Au, D. W. K., AND W. L. PERRYMAN 1985. Dolphin habitats in the eastern tropical Pacific. Fishery Bulletin, U.S. 83:623-643. BAKER, A. N. 1983. Whales and dolphins of New Zealand; an identification guide. Victoria University Press, Wellington. 133 pp. BARBER, R. T., AND F. P. CHAVEZ. 1983. Biological consequences of El Niño. Science [AD] 222:1203-1210. GASKIN, D. E. 1982. The ecology of whales and dolphins, Heinemen, London and Exeter, New Hampshire. 459 pp. HEYNING, J. E. 1984. Functional morphology involved in intraspecific fighting of the [AD] beaked whale, Mesoplodon carlhubbst. Canadian Journal of Zoology 62:1645-1654. LEATHER WOOD, S., R. R. REEVES AND L. FOSTER. 1983. The Sierra Club handbook of whales and dolphins. Sierra Club Books, San Francisco. 302 pp. MCGOWAN, J. A. 1972. The nature of oceanic ecosystems Pages 9-28 in C. B. Miller, ed. The biology of the oceanic Pacific. Proceedings of the 33rd Annual Biology Colloquium, Oregon State University Press, Corvallis, Oregon. MEAD, J. G. 1981. First records of Mesoplodon hectori (Ziphiidae) from the northern hemisphere and a description of the adult male. Journal of Mammalogy 62:430- 432. MEAD, J. G. 1984. Survey of reproductive data for the beaked whales (Ziphiidae). Pages 91-96 in W. F. Perrin et al., eds. Cetacean reproduction: estimating parameters [PAGE BREAK] 350 MARINE MAMMAL SCIENCE, VOL 3, NO. 4, 1987 only to sightings with adult males present. For 16 sightings where color morph information was obtained for all of the animals present, 5 were lone males, 6 were male/female (or possibly male/juvenile) pairs, and 5 involved single males with 2 or more female/juveniles, including one sighting with an estimated 8 animals. At sea, this whale behaves similarly to other Mesoplodon species. It is usually seen rolling over slowly at the surface or traveling at a moderate pace and in fairly tight groups when more than one animal is present. It generally does not engage in any sort of "playful" activity, e.g., breaching, tail-slapping, or spy-hopping, though on one occasion a lone male was seen to breach clear of the water three times in a row. Like other members of the genus, its behavior repertoire, at least when it is at the surface, appears to be very limited and stereotyped. Under normal circumstances there is no visible blow. IDENTITY OF THE WHALE In considering the possibilities for identification, two genera of beaked whales, Berardius and Hyperoodon, can be eliminated because of excessive size (approx- imately 12 and 7 m, respectively). Ziphius cavirostris is tan or whitish with a stubby beak and is a species we commonly identify in the eastern tropical Pacific (Pitman, unpublished observations). Tasmacetus shepherdi is probably too large also (maximum length at least 7 m, Mead 1984) and its distribution appears to be circumpolar in southern oceans (Gaskin 1982). The relatively small size (5-5.5 m), distinct beak, and small but discernable melon lead us to identify the whale with the genus Mesoplodon. Currently there are 12 recognized living species of Mesoplodon. From descrip- tions and photographs of stranded animals, the external pigmentation patterns of most of these have gradually become known (e.g., Baker 1983, Leatherwood et al. 1983), and none matches the description of the unidentified whale. (A possible exception to this, M. (Indopacetus) pacificus, is discussed below.) Nearly all of the information on Mesoplodon distribution has come from stranded specimens. Gaskin (1982) stated that the genus "seems to be in a process of prolific speciation, possibly as a result of population isolation events which began in the Pliocene and were completed in the Pleistocene." Given the large number of species in this open ocean group, it is not surprising that the distributions of individual species appear to be limited to distinct water masses (Mead 1981, Mead et al. 1982). Of the species known to inhabit tropical or warm temperate waters, M. europaeus is known only from the Atlantic, M. gingkodens inhabits the western Pacific (though apparently strays to the eastern Pacific), and M. densirostris is pantropical. One of us (RLP) has identified M. densirostris several times in the eastern tropical Pacific but only west of the area where the unidentified whale occurs. To date, there has not been any Mesoplodon specimen material collected from within the area that appears to be the normal range of the unidentified whale (J. Mead, personal communication). Tropical surface water and the apparent range of the unidentified whale (Fig. 1) both correspond to a distinct and highly productive oceanographic area that forms a uni McGowan is also near longirostris) eastern trof A likely that eastern troph Possibili 1) It m 2) Mesi wha only nort extr areas 3) It r The id collected. We wo Q y capable as Perrin ima L. López e acknowle Au, D. Pac BAKER, Vic BARBER, 22 GASKIN, Ex- HEYNIN be LEATHEI W McGo MEAD, ec C h 41 MEAD, F [PAGE BREAK] 348 MARINE MAMMAL SCIENCE, VOL. 3, NO. 4, 1987 On the black and white morph, a broad white or cream-colored swathe originates immediately posterior to the dorsal surface of the head and runs posterio-ventrally on either side of the animal. Anteriorly, the swathe is rather broad, maybe 60-90 cm, but appears to taper out toward the trailing end. This swathe has been seen to form a very large and contrasting chevron when the animal is viewed head on (and probably from above also). When seen at a distance, the posterior edge of this chevron forms a fairly distinct, contrasting border against the dark color of the rest of the posterio-dorsal surface of the whale. On closer inspection however, this border is seen to be broken up by what appears to be white dappling on a dark background. A possible clue concerning the ontogeny of the chevron pattern came from a sighting of an animal, apparently a young adult, that had the contrasting white chevron pattern described above, but it was formed entirely of individual white spots. The white spotting may become confluent as the animal matures, eventually forming the characteristic chevron pattern. The head and beak area of the black and white animal is darker than the chevron but lighter than the back and usually appears reddish brown or tan (Fig. 2). Whenever the black and white animal has been seen at relatively close range, scratches and scarring have been noted, particularly along the trailing edge of the chevron (see Fig. 2). No scars have been seen on the lighter brown animals. In most species of Mesoplodon, only adult males have teeth that erupt from the surface of the gum (usually one on either side of the lower jaw). These teeth generally protrude as tusks and, because of intraspecific fighting, males scar each other up considerably at times. Consequently, adult males are often discernable from females and young by the presence of extensive scarring (Heyning 1984). The prevalence of scarring on the black and white animals suggests that the unidentified beaked whale is sexually dimorphic and that the larger, more conspicuously marked black and white animals are adult males, while the smaller, browner, unscarred animals are females and young. The second, ostensibly female, color form appears uniform gray/brown or bronze-colored. No conspicuous color pattern, markings or scarring have been noticed on this form. The dorsal fin of the brown animal is also low, wide- based and triangular, although it tends to be slightly more falcate than in the males. The brown animals are relatively nondescript and separating them from other mesoplodonts in the field may not be possible. In fact, all of the positive sightings shown in Figure 1 were based on the presence of at least one readily identifiable adult male of the species. Mead et al. (1982) described a similar sexual dimorphism with respect to external color patterns in M. carlhubbsi. In that species, the adult male has a rostrum and a "distinctive white patch centered on the "brilliant white' dorsal prominence of the melon," while the remainder of the body is dark gray or black. Female and subadult M. carlhubbsi show an incipient version of this pattern but are more uniformly gray-brown. Figure 3 is an artist's rendition of both morphs of the unidentified whale based on photographs (Fig. 2), and descriptions and sketches made during field observations. The ventral color patterns shown are inferential, as that part of S S O Figure 3. 2 and illustr male, while b juveniles (set and the ven y e the animal has not be lodont hea species. No eru specifically do, they e carlhubbs DISTRIBU The n eastern t Wyrtki warm w ≥27°C were as sighting anomal (Barber BIOLOC Her 2 or 3 = A th [PAGE BREAK] PITMAN ET AL.: UNIDENTIFIED BEAKED WHALE 349 Figure 3. An artist's depiction of the unidentified beaked whale drawn from Figure 2 and illustrations and descriptions made in the field. The larger animal is a presumed male, while the smaller animal (not to scale) shows the pattern of presumed female/ juveniles (see text). We have seen little more of this animal than is visible in Figure 2 and the ventral patterns shown here are inferred. Illustration by Lou Silva. the animal has never been seen. Likewise, the shape of the beak and line has not been adequately examined; the illustration shows a generalized mesop- lodont head and rostrum of about the right proportions for the unidentified species. gape No erupted teeth have been seen on this species either, though we have specifically looked for them. We suspect if they exist, and they almost certainly do, they are probably not as large or conspicuous as in M. layardii or even M. carlhubbsi. A [2 S S b DISTRIBUTION The normal range of the unidentified whale appears to be restricted to the eastern tropical Pacific (Fig. 1). Nearly all of our sightings were within an area Wyrtki (1967) characterized as tropical surface water. This is an area of very warm water and recorded sea surface temperatures for 20 of our sightings were ≥27°C (80°F). The two sightings in Figure 1 that appear south of this range were associated with southerly intrusions of warm water. The southernmost sighting was made in February 1983 when a major El Niño event was shifting anomalously warm water and associated organisms south along the coast of Peru (Barber and Chavez 1983, Velez et al. 1984). BIOLOGICAL OBSERVATIONS Herd size for the 24 positive sightings ranged from 1 to 8, with groups of 2 or 3 being the most common. As noted above however, these results pertain a [PAGE BREAK] 352 MARINE MAMMAL SCIENCE, VOL. 3, NO. 4, 1987 for stock assessment and management. Reports of the International Whaling Com- mission, Special Issue 6. MEAD, J. G., W. A. WALKER AND W. J. HOUCK. 1982. Biological observations on Mesoplodon carlhubbsi (Cetacea. Ziphiidae). Smithsonian Contributions to Zoology 344. 25 pp. PERRIN, W. F., M. D. SCOTT, G. J. WALKER, F. M. RALSTON AND D. W. K. Au. 1983. Distribution of four dolphins (Stenella spp. and Delphinus delphis) in the eastern tropical Pacific, with an annotated catalogue of data sources. NOAA Technical Memorandum NMFS-SWFC-38. 65 PP. PERRIN, W. F., M. D. SCOTT, G. J. WALKER AND V. L. CASS. 1985. Review of geographical stocks of tropical dolphins (Stenella spp. and Delphinus delphis) in the eastern tropical Pacific. NOAA Technical Report NMFS 28. 28 pp. REID, J. L., JR. 1962. On circulation, phosphate-phosphorus content, and zooplankton volumes in the upper part of the Pacific. Limnology and Oceanography 7:287-306. VELEZ, J., J. ZEBALLOS, AND M. MENDEZ. 1984. Effects of the 1982-83 El Niño on the fishes and crustaceans off Peru. Tropical Ocean-Atmosphere Newsletter 28: 10-12. WYRTKI, K. 1967. Circulation and water masses in the eastern equatorial Pacific Ocean. International Journal of Oceanology and Limnology 1:117-147. MARINE MA © 1987 by th BEH Sightings K. breviceps what is kno Received: March 7, 1986 Accepted: May 11, 1987 at sea (Yam whale (Rob A review tuna purse noteworthy 11°17'N, calf, were s albacares, a and spinne based on s as they did during the While i material, released th to hide he an area o reported F instances During minutes a and then off the ne out" of rammed then bot whales w pugnacio whale (F These [PAGE BREAK] MARINE MAMMAL SCIENCE, 3(4):345-352 (October 1987) 1987 by the Society for Marine Mammalogy OBSERVATIONS OF AN UNIDENTIFIED BEAKED WHALE (MESOPLODON SP.) IN THE EASTERN TROPICAL PACIFIC ROBERT L. PITMAN Southwest Fisheries Center, [AD] P.O. Box 271, La Jolla, California 92038 ANELIO AGUAYO L. JORGY URN R. Facultad de Ciencias, UNAM, Apartado Postal 70 572, CP. 04510, México, DF, México ABSTRACT Multiple sightings of a distinctive but unidentified species of beaked whale have been made in the eastern tropical Pacific. The unidentified whale has two color morphs: a conspicuously marked black and white form (judged to be larger), and a uniformly gray-brown form Maximum length estimates have been 5-5.5 m. Other features include a relatively flat head, with a small, distinct melon; a moderately long beak; and a low, wide-based, triangular dorsal fin. On most animals the trailing edge of the dorsal fin is only slightly falcate and often appears straight. On the black and white morph, a broad white or cream-colored swathe originates immediately posterior to the dorsal surface of the head and runs posterio-ventrally on either side of the animal. The prevalence of scarring on the black and white animals suggests sexual dimorphism and that these larger, more conspicuously marked animals are adult males, while the smaller, browner, unscarred animals are females and young. Possibilities for identification include: 1) a well-marked race of a known Mesoplodon sp., 2) Mesoplodon (Indopacetus) pacificus or 3) an undescribed species. Key words: Mesoplodon, unidentified beaked whale, eastern tropical Pacific. In this paper we report on multiple sightings of a distinctive but as yet unidentified species of beaked whale from the eastern tropical Pacific (ETP). A morph of this whale that we assume is the adult male (see below) is conspicuously marked and eminently identifiable in the field. Despite this, it does not fit the description of any known species of ziphiid. We provide here a field description of the whale, identify its range and comment on its identity. 345 [PAGE BREAK] 346 MARINE MAMMAL SCIENCE, VOL. 3, NO. 4, 1987 30° N Hawallan Islands ⚫ Subtropical 20° Seasonally Tropical 10° Tropical. Equatorial 10° 20°S 160° 150° Subtropical 140° 130° 120° 110° Mexico 100° O Sightings Tentative sightings Survey vessel noon positions (soo text) 90° 80° South America 70° Figure 1. Sightings of the unidentified beaked whale in the eastern tropical Pacific relative to surface water types (after Wyrtki 1967). PITMAN ET AL.: UNIDENTIFIED BEAKED WHALE 347 a SIGHTINGS Figure 1 shows 24 positive and 8 tentative sightings of the unidentified whale. All except one were made by trained observers during marine mammal survey cruises for the Southwest Fisheries Center in the central and eastern tropical Pacific (see Perrin et al. 1983). Noon positions of survey vessels during these cruises are also plotted in Figure 1 and indicate the extent of survey coverage. The remaining sighting (Fig. 2) was photographed during a marine mammal identification course taught by two of us (AAL and JUR) at Facultad de Ciencias, Universidad Nacional Autonoma de Mexico (UNAM). Figure 2. b Photographs of the unidentified beaked whale from Bahia de Banderas, Jalisco, Mexico. The animal shown in 2a and 2b, foreground, is presumably an adult male; notice the scratches behind the white area. Photos by P. Hernández and J. L. López. DESCRIPTION The unidentified whale has two distinct color morphs: a conspicuously marked black and white form and a relatively nondescript morph that appears to be uniformly gray-brown (Figs. 2 and 3). When they have been seen together, the black and white animal has been judged by several observers to be the larger of the two. Estimates of maximum length have most often been between approximately 5-5.5 m, making it an average-to-slightly-large-sized Mesoplodon (see Mead 1984). The head is relatively flat, as in most mesoplodonts, with a small but distinct melon. The beak is moderately long, appearing similar in size to that of M. densirostris. On one occasion an observer reported that the mandible was paler than the upper part of the beak on one of the black and white animals. The distinctive dorsal fin is low, wide-based and triangular. On most animals the trailing edge of the fin is only slightly falcate and often appears straight. Most of the dorsal surface of the black and white form, including the dorsal fin, is dark and has been variously described as blackish, black/brown, chocolate brown or dark olive brown. This is similar to the range in color variation noted for short-finned pilot whales (Globicephala macrorhynchus) in the eastern Pacific. Pilot whales in the field are generally described as jet black, but in the ETP they often appear greenish brown. Whether these perceived differences are the result of lighting effects, as seems likely, or some actual physical differences, is not known. For the time being, the unidentified whale is best described as black and white. [PAGE BREAK] 348 MARINE MAMMAL SCIENCE, VOL. 3, NO. 4, 1987 On the black and white morph, a broad white or cream-colored swathe originates immediately posterior to the dorsal surface of the head and runs posterio-ventrally on either side of the animal. Anteriorly, the swathe is rather broad, maybe 60-90 cm, but appears to taper out toward the trailing end. This swathe has been seen to form a very large and contrasting chevron when the animal is viewed head on (and probably from above also). When seen at a distance, the posterior edge of this chevron forms a fairly distinct, contrasting border against the dark color of the rest of the posterio-dorsal surface of the whale. On closer inspection however, this border is seen to be broken up by what appears to be white dappling on a dark background. A possible clue of the chevron pattern came from a sighting of an concerning the ontogeny animal, apparently a young adult, that had the contrasting white chevron patter described above, but it was formed entirely of individual white spots. The whi spotting may become confluent as the animal matures, eventually forming th characteristic chevron pattern. The head and beak area of the black and whi animal is darker than the chevron but lighter than the back and usually appears reddish brown or tan (Fig. 2). Whenever the black and white animal has been seen at relatively close range, scratches and scarring have been noted, particularly along the trailing edge of the chevron (see Fig. 2). No scars have been seen on the lighter brown animals. In most species of Mesoplodon, only adult males have teeth that erupt from the surface of the gum (usually one on either side of the lower jaw). These teeth generally protrude as tusks and, because of intraspecific fighting, males scar each other up considerably at times. Consequently, adult males are often discernable from females and young by the presence of extensive scarring (Heyning 1984). The prevalence of scarring on the black and white animals suggests that the unidentified beaked whale is sexually dimorphic and that the larger, more conspicuously marked black and white animals are adult males, while the smaller, browner, unscarred animals are females and young. The second, ostensibly female, color form appears uniform gray/brown or bronze-colored. No conspicuous color pattern, markings or scarring have been noticed on this form. The dorsal fin of the brown animal is also low, wide- based and triangular, although it tends to be slightly more falcate than in the males. The brown animals are relatively nondescript and separating them from other mesoplodonts in the field may not be possible. In fact, all of the positive sightings shown in Figure 1 were based on the presence of at least one readily identifiable adult male of the species. Mead et al. (1982) described a similar sexual dimorphism with respect to external color patterns in M. carlhubbsi. In that species, the adult male has a "brilliant white' rostrum and a "distinctive white patch centered on the dorsal prominence of the melon," while the remainder of the body is dark gray or black. Female and subadult M. carlhubbsi show an incipient version of this pattern but are more uniformly gray-brown. Figure 3 is an artist's rendition of both morphs of the unidentified whale based on photographs (Fig. 2), and descriptions and sketches made during field observations. The ventral color patterns shown are inferential, as that part of Figure 3. PITMAN ET AL.: UNIDENTIFIED BEAKED WHALE 349 An artist's depiction of the unidentified beaked whale drawn from Figure 2 and illustrations and descriptions made in the field. The larger animal is a presumed male, while the smaller animal (not to scale) shows the pattern of presumed female/ juveniles (see text). We have seen little more of this animal than is visible in Figure 2 and the ventral patterns shown here are inferred. Illustration by Lou Silva. the animal has never been seen. Likewise, the shape of the beak and gape line has not been adequately examined; the illustration shows a generalized mesop- lodont head and rostrum of about the right proportions for the unidentified species. No erupted teeth have been seen on this species either, though we have specifically looked for them. We suspect if they exist, and they almost certainly do, they are probably not as large or conspicuous as in M. layardii or even M. carlhubbsi. DISTRIBUTION The normal range of the unidentified whale appears to be restricted to the eastern tropical Pacific (Fig. 1). Nearly all of our sightings were within an area Wyrtki (1967) characterized as tropical surface water. This is an area of very warm water and recorded sea surface temperatures for 20 of our sightings were ≥27°C (80°F). The two sightings in Figure 1 that appear south of this range were associated with southerly intrusions of warm water. The southernmost sighting was made in February 1983 when a major El Niño event was shifting anomalously warm water and associated organisms south along the coast of Peru (Barber and Chavez 1983, Velez et al. 1984). BIOLOGICAL OBSERVATIONS Herd size for the 24 positive sightings ranged from 1 to 8, with groups of 2 or 3 being the most common. As noted above however, these results pertain [PAGE BREAK] 350 MARINE MAMMAL SCIENCE, VOL. 3, NO. 4, 1987 only to sightings with adult males present. For 16 sightings where color morph information was obtained for all of the animals present, 5 were lone males, 6 were male/female (or possibly male/juvenile) pairs, and 5 involved single males with 2 or more female/juveniles, including one sighting with an estimated 8 animals. At sea, this whale behaves similarly to other Mesoplodon species. It is usually seen rolling over slowly at the surface or traveling at a moderate pace and in fairly tight groups when more than one animal is present. It generally does not engage in any sort of "playful" activity, e.g., breaching, tail-slapping, or spy-hopping, though on one occasion a lone male was seen to breach clear of the water three times in a row. Like other members of the genus, its Lavior repertoire, at least when it is at the surface, appears to be very limand stereotyped. Under normal circumstances there is no visible blow. IDENTITY OF THE WHALE In considering the possibilities for identification, two genera of beaked whales, Berardius and Hyperoodon, can be eliminated because of excessive size (approx- imately 12 and 7 m, respectively). Ziphius cavirostris is tan or whitish with a stubby beak and is a species we commonly identify in the eastern tropical Pacific (Pitman, unpublished observations). Tasmacetus shepherdi is probably too large also (maximum length at least 7 m, Mead 1984) and its distribution appears to be circumpolar in southern oceans (Gaskin 1982). The relatively small size (5-5.5 m), distinct beak, and small but discernable melon lead us to identify the whale with the genus Mesoplodon. Currently there are 12 recognized living species of Mesoplodon. From descrip- tions and photographs of stranded animals, the external pigmentation patterns of most of these have gradually become known (e.g., Baker 1983, Leatherwood et al. 1983), and none matches the description of the unidentified whale. (A possible exception to this, M. (Indopacetus) pacificus, is discussed below.) Nearly all of the information on Mesoplodon distribution has come from stranded specimens. Gaskin (1982) stated that the genus "seems to be in a process of prolific speciation, possibly as a result of population isolation events which began in the Pliocene and were completed in the Pleistocene." Given the large number of species in this open ocean group, it is not surprising that the distributions of individual species appear to be limited to distinct water masses (Mead 1981, Mead et al. 1982). Of the species known to inhabit tropical or warm temperate waters, M. europaeus is known only from the Atlantic, M. gingkodens inhabits the western Pacific (though apparently strays to the eastern Pacific), and M. densirostris is pantropical. One of us (RLP) has identified M. densirostris several times in the eastern tropical Pacific but only west of the area where the unidentified whale occurs. To date, there has not been any Mesoplodon specimen material collected from within the area that appears to be the normal range of the unidentified whale (J. Mead, personal communication). Tropical surface water and the apparent range of the unidentified whale (Fig. 1) both correspond to a distinct and highly productive oceanographic area that PITMAN ET AL. UNIDENTIFIED BEAKED WHALE 351 forms a unique biotic province in the eastern Pacific (Reid 1962, Wyrtki 1967, McGowan 1972, Au and Perryman 1985). The range of the unidentified whale is also nearly identical to the range of the eastern spinner dolphin (Stenella longirostris), a pantropical species with a well-marked form endemic to the eastern tropical Pacific (Perrin et al. 1985). From the evidence above, it seems likely that a distinct, possibly endemic, species of Mesoplodon also occurs in the eastern tropical Pacific. Possibilities for identification of the unidentified whale include the following: 1) It may be a well-marked race of a known species of Mesoplodon. 2) Mesoplodon (Indopacetus) pacificus is currently the rarest extant species of whale known and has never been identified in the flesh. It is known from only two skulls: one from Queensland, Australia, and one from Somalia, northeast Africa. It is possible, however, that these records represent extralimital strays from a localized population inhabiting a little-explored area, such as the eastern tropical Pacific. 3) It may be an undescribed species. The identity of the whale will remain unknown until a stranded specimen is collected. ACKNOWLEDGMENTS We would like to thank Jim Cotton, Gary Friedrichsen and Scott Sinclair for their capable assistance in the field. David Au, Jim Mead, John Heyning and especially William Perrin improved the manuscript with their comments. We thank P. Hernández and J. L. López for use of their photographs, Henry Orr for drafting Figure 1, and we gratefully acknowledge the talents of Lou Silva in preparing Figure 3. LITERATURE CITED Au, D. W. K., AND W. L. PERRYMAN 1985. Dolphin habitats in the eastern tropical Pacific. Fishery Bulletin, U.S. 83:623-643. BAKER, A. N. 1983. Whales and dolphins of New Zealand; an identification guide. Victoria University Press, Wellington. 133 pp. BARBER, R. T., AND F. P. CHAVEZ. 1983. Biological consequences of El Niño. Science [AD] 222;1203-1210. GASKIN, D. E. 1982. The ecology of whales and dolphins, Heinemen, London and Exeter, New Hampshire. 459 pp. HEYNING, J. E. 1984. Functional morphology involved in intraspecific fighting of the [AD] beaked whale, Mesoplodon carlhubbsi. Canadian Journal of Zoology 62:1645-1654. LEATHERWOOD, S., R. R. REEVES AND L. FOSTER. 1983. The Sierra Club handbook of whales and dolphins. Sierra Club Books, San Francisco. 302 pp. MCGOWAN, J. A. 1972. The nature of oceanic ecosystems. Pages 9-28 in C. B. Miller, ed. The biology of the oceanic Pacific. Proceedings of the 33rd Annual Biology Colloquium, Oregon State University Press, Corvallis, Oregon. MEAD, J. G. 1981. First records of Mesoplodon hectori (Ziphiidae) from the northern hemisphere and a description of the adult male. Journal of Mammalogy 62:430- 432. MEAD, J. G. 1984. Survey of reproductive data for the beaked whales (Ziphiidae). Pages 91-96 in W. F. Perrin et al., eds. Cetacean reproduction: estimating parameters [PAGE BREAK] 352 MARINE MAMMAL SCIENCE, VOL. 3, NO. 4, 1987 for stock assessment and management. Reports of the International Whaling Com- mission, Special Issue 6. MEAD, J. G., W. A. WALKER AND W. J. HOUCK. 1982. Biological bservations on Mesoplodon carlhubbsi (Cetacea: Ziphiidae). Smithsonian Contributors to Zoology 344. 25 PP- Au. 1983. PERRIN, W. F., M. D. SCOTT, G. J. WALKER, F. M. RALSTON AND D. Distribution of four dolphins (Stenella spp. and Delphinus delp in the eastern tropical Pacific, with an annotated catalogue of data sources. NA Technical Memorandum NMFS-SWFC-38. 65 pp. PERRIN, W. F., M. D. SCOTT, G. J. WALKER AND V. L. CASS. 1985 Review of geographical stocks of tropical dolphins (Stenella spp. and Delphinus delphis) in the eastern tropical Pacific. NOAA Technical Report NMFS 28. 28 pp. REID, J. L., JR. 1962. On circulation, phosphate-phosphorus content, and zooplankton volumes in the upper part of the Pacific. Limnology and Oceanography 7:287–306. VELEZ, J., J. ZEBALLOS, AND M. MENDEZ. 1984. Effects of the 1982-83 El Niño on the fishes and crustaceans off Peru. Tropical Ocean-Atmosphere Newsletter 28: 10-12. WYRTKI, K. 1967. Circulation and water masses in the eastern equatorial Pacific Ocean. International Journal of Oceanology and Limnology 1.117-147. Received: March 7, 1986 Accepted: May 11, 1987 [PAGE BREAK] [17 decembre 1892, détart de Bransfield, ocean antarctique, su le Balaena FROM EDINBURGH TO THE ANTARCTIC 209 Just after killing the seal there was a shout amongst the men forward, 'A Uni! A Uni!'-the whalers' term for a Narwhale. Several men said they saw their horns. de unicorn. évidemment The crow's-nest cask, about five was sent aloft to-day. It is a feet deep, paint- ed white, with iron clamps that clasp on to the main-topgallant mast. In the bottom there is a trap-door. To get into the nest you climb up a Jacob's ladder- wooden ratlins rigged on two backstays that run from the top-gallant mast-head to the cross-trees; these run through the bottom of the tub. You climb up these and shove the trap open with your head, and when you are right into the tub you let the trap shut below you, and stand on it, and enjoy the extensive view. If you prefer it, you can sit on a shelf-seat fixed in the back of the tub-a sheltered, quiet place, far removed from the troubles of the little world below: round the top of the tub there is a small iron balustrade, on which a screen runs, so as to shelter the watcher from the wind. The boats were all lowered from the skids to the MUSÉE OCEANOGRAPHIQUE DE MONACO [PAGE BREAK] MARINE MAMMAL SCIENCE, 18(3):577-608 (July 2002) ©2002 by the Society for Marine Mammalogy A NEW SPECIES OF BEAKED WHALE MESOPLODON PERRINI SP. N. (CETACEA: ZIPHIIDAE) DISCOVERED THROUGH PHYLOGENETIC ANALYSES OF MITOCHONDRIAL DNA SEQUENCES MEREL L. DALEBOUT School of Biological Sciences, University of Auckland, Private Bag 92019, Auckland 1000, New Zealand E-mail: m.dalebout@auckland.ac.nz JAMES G. MEAD National Museum of Natural History, Mailstop NHB 108, Smithsonian Institution, Washington, DC 20560, U.S.A. C. SCOTT BAKER' School of Biological Sciences, University of Auckland, Private Bag 92019, Auckland 1000, New Zealand ALAN N. BAKER Science & Research Unit, Department of Conservation, P. O. Box 10-420, Wellington, New Zealand ANTON L. VAN HELDEN Museum of New Zealand Te Papa Tongarewa, P. O. Box 467, Wellington, New Zealand ABSTRACT Mesoplodon perrini, a new species of beaked whale is described on the basis of five animals stranded on the coast of California (between 32°55'N, 117°15'W and 36°37'N, 121°55'W) from May 1975 to September 1997. Four of these animals were initially identified as Hector's beaked whales M. bectori based on cranial morphology (Mead 1981). A fifth specimen was ini- tially identified as a neonate Cuvier's beaked whale Ziphius cavirostris based on external features. These specimens were first recognized as representatives of an undescribed species through phylogenetic analysis of mitochondrial (mt) DNA control region and cytochrome b sequence data. Although similar mor- phologically, the genetic data do not support a close evolutionary relationship between M. perrini and M. hectori. Instead, these data suggest a possible sister- 'Corresponding author; e-mail: cs.baker@auckland.ac.nz. 577 [PAGE BREAK] 578 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 species relationship with the lesser beaked whale M. peruvianus. Sightings of two small beaked whales off California in the 1970s which were tentatively identified as M. hectori are also likely to be M. perrini. We suggest that M. hectori is confined to the Southern Hemisphere, while M. perrini is known to date only from the North Pacific. Key words: molecular genetics and systematics, morphology, external appear- ance, natural history, distribution, Mesoplodon perrini, Perrin's beaked whale. Beaked whales (Cetacea: Ziphiidae) are the least known of all cetacean fam- ilies. In terms of species diversity, they are second only to oceanic dolphins (Family Delphinidae), with 20 species currently recognized. Beaked whales are rarely observed at sea due to their preference for deep ocean waters and elusive habits. Most species are known from only a small number of stranded speci- mens, and several have never been seen alive. Of the twelve cetacean species described in the last 100 years, eight have been ziphiids, primarily of the genus Mesoplodon. In the closing decade of the 20th century, two new beaked whales were discovered; the lesser beaked whale M. peruvianus (Reyes et al. 1991), and Bahamonde's beaked whale M. bahamondi (Reyes et al. 1995), although the latter is now recognized as synonymous with M. traversii (Gray, 1874) (van Helden et al. 2002). Here we document the occurrence and characteristics of a previously un- described species of Mesoplodon beaked whale in the North Pacific Ocean. In the mid to late 1970s, four beaked whales stranded within 85 km of each other along the southern coast of California (Table 1). These animals were identified as Hector's beaked whales M. hectori, the first and only records of this species from the Northern Hemisphere (Mead 1981). In 1997 a database of mitochondrial (mt) DNA control region reference sequences was compiled to assist in beaked whale species identification (Dalebout et al. 1998, Henshaw et al. 1997). All specimens in this reference database were validated through examination by experts in cetacean morphology and the collection of diag- nostic skeletal material or photographic records (Dizon et al. 2000). Among the specimens incorporated in these analyses was one of the "M. hectori" from California (USNM504259; Henshaw et al. 1997). A phylogenetic review of the database, which at this time consisted of reference sequences from 16 of the 20 described species, including Southern Hemisphere M. hectori, suggested that the California specimen was not of this species nor any other species in the database (Dalebout et al. 1998). To further investigate this anomaly, DNA was extracted from the remaining three specimens from California described by Mead (1981). Phylogenetic anal- yses of mtDNA control region and cytochrome b sequences from these spec- imens, using a complete reference database which now includes all 20 de- scribed beaked whale species, confirmed that all four were of the same species, yet did not represent M. hectori nor any other known ziphiid species (Dalebout 2002). Instead, these results suggested that the California specimens repre- sented an undescribed species of beaked whale. Here we present a formal description of this new species, and include a description of a fifth specimen, [PAGE BREAK] Table 1. Specimens of Mesoplodon perrini from California, in order by stranding date. Total length in cm. DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE 579 Museum number Field number Date found USNM504259 USNM504260 USNM504853 LAM088901 JRH 052 TMMC-C75 1975 May 22 1975 May 28 1978 Sept 9 1979 Dec 27 Locality Camp Pendleton Coordinates Camp Pendleton Carlsbad Total length 33°15'N, 117°26'W 210 33°16'N, 117°26'W 443 33°07'N, 117°20'W 390 Sex M F M 1997 Sept 18 Torrey Pines State Reserve Monterey 32°55'N, 117°15'W 36°37'N, 121°55'W 245 224 M M [PAGE BREAK] 580 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 which stranded in Monterey, California, in 1997, and was initially identified as a neonate Cuvier's beaked whale Ziphius cavirostris from external morphol- ogy. METHODS Material Examined Five specimens of the undescribed species from California were examined (Table 1). Phylogenetic analyses of mtDNA control region and cytochrome b ziphiid species. sequences were used to compare these specimens to all 20 previously described Museums and institutions holding specimens of the new species are as fol- lows: Los Angeles County Museum of Natural History, California (LAM) and the National Museum of Natural History, Smithsonian Institution, Washing- ton, DC (USNM). Tissue samples are held by: the School of Biological Sci- ences, University of Auckland, New Zealand (AUNZ); Southwest Fisheries Science Centre, La Jolla, CA (SWFSC); and, The Marine Mammal Centre, Sausalito, CA (TMMC). Morphological comparisons were made to 12 specimens of M. hectori held (NMNZ), 11 = by the following museums: Museum of New Zealand Te Papa Tongarewa 7; The Australian Museum, Sydney (AMS), " 1; Tasmanian Museum and Art Gallery (TAM), n 1; the South Australian Museum (SAM), n = 1; and, Museo Acatushún de Aves y Mamíferos Australes (MAAMA), Tierra del Fuego, n = 2. Further information on specimens of M. hectori ex- amined can be obtained from JGM or AVH. DNA Extraction and Sequencing DNA was extracted from the teeth and cartilage on the museum-held ma- terial of the four specimens (USNM504259, USNM504260, USNM504853, LAM088901) described by Mead (1981), using the silica-based method of Höss and Pääbo (1993), as modified by Matisoo-Smith et al. (1997), and techniques described in Pichler et al. (2001). Museum material from USNM504259 was included to confirm that the original soft tissue sample (SWFSC-24976) used by Henshaw et al. (1997) had indeed been derived from this specimen. (Soft tissue samples were not available for the other three spec- imens). Total genomic DNA was extracted from soft tissue samples obtained from the Monterey specimen (TMMC-C75) using standard methods (Sam- brook et al. 1989), as modified by Baker et al. (1993). Samples were stored in salt-saturated, dimethylsulphoxide (DMSO) solution prior to analysis. Segments of the 5' end of the mtDNA control region and 5' end of the cytochrome b gene were amplified and sequenced from all five specimens, and aligned to the sequences already in the beaked whale reference database, as described in Dalebout (2002). [PAGE BREAK] DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE Phylogenetic Analyses 581 Phylogenetic relationships among the California specimens and the 20 known beaked whale species were reconstructed from sequence data from both mtDNA loci using maximum likelihood (ML) methods (consensus length of alignments, control region, 437 bp; cytochrome b, 384 bp; Dalebout 2002). Baird's beaked whale, Berardius bairdii, which likely represents the most basal extant species in this family (e.g., Dalebout et al. 1998) was used as an out- group. To further investigate sister-species relationships among a subset of Meso- plodon species, including the California specimens, the control region and cy- tochrome b sequences were combined to increase phylogenetic signal (consen- sus length of alignment, 821 bp). A partition-homogeneity test (Farris et al. 1995) indicated that these loci were congruent (branch and bound search, 1,000 replicates). Although the individuals representing each species differed between the two loci in some cases, this was not considered a problem as intraspecific variation was generally much lower than interspecific variation (see discussion below). Starting parameters for ML reconstruction were esti- mated from an initial neighbour-joining tree built using general-time-revers- ible (GTR) distances. For ML analyses, the heuristic search option, with ran- dom sequence addition (100 replicates), and sub-tree pruning-regrafting branch swapping, was used. The statistical consistency of groupings was eval- uated by 200 ML bootstrap resamplings of the data. Bremer support was calculated using Tree Rot v.2a (Sorensen 1999), based on one of the three most parsimonious trees (= ML tree) obtained through an exhaustive parsimony search. All phylogenetic analyses were conducted using the program PAUP* 4.0 beta 6 (Swofford 1999). RESULTS DNA Sequence Data For the mtDNA control region, fragments ranging in length from 245 bp to 434 bp were sequenced successfully from the five California specimens. For the mtDNA cytochrome b, fragments ranging in length from 276 bp to 384 bp were sequenced successfully. For those specimens represented by hard tissue (i.e., tooth or cartilage; USNM504260, USNM504853, LAM088901), only shorter fragments were obtained, as expected from DNA extractions from such material (Höss and Pääbo 1993). Comparison of the mtDNA control region sequence published by Henshaw et al. (1997) (Genbank Accession No. U70466) with that obtained from museum-held material from USNM504259 confirmed that they were identical, and as such likely derived from the same specimen. All previously unpublished sequences have been deposited in Gen- bank (Accession No.'s: AF441254-AF441263). [PAGE BREAK] 582 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 Phylogenetic Analyses Phylogenetic analyses of sequence data from the mtDNA control region (Fig. 1) and cytochrome b (not shown) confirmed that all five California spec- imens were of the same species, and distinct from all other 20 known species of beaked whale. Although higher-level relationships were not well resolved by these rapidly evolving mtDNA loci (i.e., bootstrap scores for most internal nodes, <50%), all species-specific groupings were supported by high bootstrap scores (>80%). MtDNA control region sequences representing the southern bottlenose whale Hyperoodon planifrons were the exception to this trend. To further investigate the genetic distinctiveness of the California animals, and exclude the possibility that they represent only a highly divergent, geo- graphic subdivision of M. hector, we compared the combined control region and cytochrome b sequences (821 bp) to a subset of potential sister-taxa (Fig. 1, gray box). While a monophyletic grouping of the California animals and M. hectori would argue for a single-species classification despite a deep diver- gence (Lento et al. 1997, Wayne et al. 1990), the results indicated instead that the California specimens were more closely related to at least four other species of Mesoplodon, than to the morphologically similar M. hectori (bootstrap score, 83%; Bremer support = 7; Fig. 2). No synapomorphies were found to unite the California specimens and M. hectori exclusive of other beaked whales. These results allowed us to reject the hypothesis of a deep intraspecific divergence, and argue instead that these specimens represent a previously unrecognized species of beaked whale. Although higher-level relationships among this subset of Mesoplodon species were not fully resolved (Fig. 2), there was some support for a sister-species relationship between the California specimens and the lesser beaked whale M. peruvianus (bootstrap score, 69%). There was also low-level support for a clade consisting of these two species, plus Gray's beaked whale M. grayi, (bootstrap score, 55%). Phylogenetic reconstructions based on nuclear sequences support a similar pattern of relationships among these species (Dalebout 2002), Intra- and Interspecific Genetic Divergence Over the 245 bp fragment of the mtDNA control region covered by se- quence data from all five specimens, all shared the same haplotype (Fig. 3). Analysis of the 280 bp fragment of the cytochrome b covered by all five specimens revealed two variable sites (one synonymous third position trans- version and one non-synonymous first position transition) defining three unique haplotypes (Fig. 4). The adult female (USNM504260), and two of the three calves (USNM504259 and TMMC-C75) share the same haplotype at this locus, while the adult male (USNM504853) and the remaining calf (LAM088901) were both unique. The adult female and the calf, USNM504259, both stranded at Camp Pendleton in the same week of May 1975 (Table 1). Comparisons of intra- and interspecific pairwise sequence divergence for all [PAGE BREAK] DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE 583 20 beaked whale species confirmed that the California specimens follow a similar pattern to other ziphiids (Fig. 5a, b). Over the 437 bp control region alignment, intraspecific variation (using two representatives per species) was found to be generally less than 2%, while interspecific variation was generally greater than 4%. Similar trends were found in a previous analysis, which compared intra- and interspecific genetic divergence among nine described beaked whale species (Dalebout et al. 1998). The new species differs from all other beaked whales by an average of 8.55% over this fragment. Over the 384 bp mtDNA cytochrome b alignment, intraspecific variation was found to be generally less than 1.5%, while interspecific variation was found to be gen- erally greater than 3% for 20 described beaked whale species. The new species differs from all other beaked whale species by an average of 15.24% over this fragment. See Dalebout (2002) for discussion regarding comparative levels of divergence at the mtDNA cytochrome b versus the control region among the Ziphiidae. DESCRIPTION Order Cetacea Brisson, 1762 Family Ziphiidae Gray, 1865 Mesoplodon perrini sp. n. HOLOTYPE Adult male (USNM504853); skull, mandible, and postcranial skeleton, at the National Museum of Natural History, Smithsonian Institution, Washing- ton, DC. This specimen was found on 9 September 1978, by G. Carsten, and collected two days later by J.G.M. TYPE LOCALITY Carlsbad, California (33°07'N, 117°20'W), United States of America. PARATYPES Male calf (USNM504259); fragmented cranium and postcranial skeleton, at the Smithsonian National Museum of Natural History, Washington, DC, col- lected by W. F. Perrin. Adult female (USNM504260); skull, mandible, and postcranial skeleton, at the Smithsonian National Museum of Natural History, Washington, DC, col- lected by W. F. Perrin. Male calf (LAM088901); skull, mandible, at the Los Angeles County Mu- seum of Natural History, collected by J. R. Henderson (JRH 052). Male calf (TMMC-C75); skull, mandible, and postcranial skeleton, at the Los Angeles County Museum of Natural History, collected by M. Haulena. [PAGE BREAK] 584 MARINE MAMMAL SCIENCE, VOL. 18, NO 3, 2002 85 100 100 H 95 Sowerby's beaked whale Mesoplodon bidens True's beaked whale Mesoplodon mirus 100 spade-toothed whale Mesoplodon traversii (=M. bahamondi) ginkgo-toothed beaked whale Mesoplodon ginkgodens Mesoplodon carlhubbsi densebeaked whale 93 95 Hubbs' beaked whale 97 94 99 Gray's beaked whale 99 Mesoplodon densirostris Stejneger's beaked whale Mesoplodon stejnegeri Mesoplodon grayi lesser beaked whale Mesoplodon peruvianus 100 Perrin's beaked whale Mesoplodon perrini sp. n. 100 straptoothed whale 58 99 Mesoplodon layardii Longman's beaked whale Indopacetus pacificus Andrews' beaked whale Mesoplodon bowdoini 84 Hector's beaked whale 83 100 98 Mesoplodon hectori Gervais' beaked whale Mesoplodon europaeus 93 northern bottlenose whale Hyperoodon ampullatus southern bottlenose whale Hyperoodon planifrons Shepherd's beaked whale Tasmacetus shepherdi Cuvier's beaked whale 96 Arnoux's beaked whale Berardius arnuxii Baird's beaked whale Berardius bairdii Ziphius cavirostris nucleotide divergence per lineage (GTR correction) 12% 8% 4% 0% Treelength=357 Consistency index (CI)=0.401 Retention index (RI)=0.712 Figure 1. Phylogenetic relationships among the 20 described species of beaked whales (Ziphiidae) from maximum-likelihood analyses, based on 437 bp of mitochon- drial DNA control region. Numbers above internal nodes indicate bootstrap values 50%. All described species represented by two reference specimens where possible. Arrows highlight respective positions of Mesoplodon perrini sp. n. and M. hectori, the species as which several specimens of this new species were initially described. Gray box indicates subset of taxa, with addition of M. hectori, used in further analyses. Note [PAGE BREAK] DALEBOUT ET AL.: NEW SPECIES OF BEAKED WHALE 585 ETYMOLOGY The specific name, perrini, was chosen as a tribute to the American cetacean biologist, Dr. W. F. Perrin, for his role in the collection of two of the known specimens of this species, and his ongoing contribution to marine mammal science and conservation. We propose this species be known by the common name, Perrin's beaked whale. DIAGNOSIS Molecular Characters M. perrini can be differentiated from all other species of Mesoplodon beaked whales based on molecular genetic characters, as demonstrated by phylogenetic analyses of mtDNA control region and cytochrome b sequences (Fig. 1-5). Over the 434 bp control region segment, M. perrini is distinguished from M. bectori by 26 diagnostic sites (5.99% pairwise sequence divergence), including two insertion-deletions (indels), and from M. peruvianus, its likely sister-spe- cies, by 16 diagnostic sites (3.46%), including one indel (Fig. 3). Over the 384 bp cytochrome b segment, M. perrini is distinguished from M. hectori by 37 diagnostic sites (9.64%), including four first position and three second position substitutions, and from M. peruvianus by 48 diagnostic sites (12.50%), including six first position and four second position substitutions (Fig. 4). In comparisons including all ziphiid species, M. perrini is distinguished by one diagnostic site (sensu Davis and Nixon, 1992) at the control region (position 111 = A; Fig. 3), and one diagnostic site at the cytochrome b (position 182 [2nd]T; Fig. 4), given a mean of two diagnostic sites per species for both fragments. Note that high levels of homoplasy were observed at these mtDNA loci due to the rapid rate of accumulation of mutations and the large number of species to be differentiated (Sanderson and Donoghue 1989). Morphological Characters The following characters of the mandibles, teeth, and skull are, when com- bined, diagnostic for M. perrini: (1) Short mandibular symphysis (19%-23% mandible length). (2) Convex profile to anterior part of mandible over the length of the sym- physis. that M. bahamondi (Reyes et al. 1995) was recently recognized as synonymous with M. traversii, a species described by J. E. Gray in 1874 (van Helden et al. 2002). The aligned sequence files used in these analyses are available electronically from http:// www.sbs.auckland.ac.nz/research-groups/ecology_and_evolution/molecular_ecol_evo l_lab or http://www.dna-surveillance.auckland.ac.nz. [PAGE BREAK] 586 MARINE MAMMAL SCIENCE, VOL 18, NO. 3, 2002 100/40 BbaSW4965/X92541 BbaBC9220 MdeNZ01/NZ02 100/18 MdeNHMUK/SW4010 Mesoplodon densirostris densebeaked whale 100/12 Mgr05/29 MgrI1/H04 Mesoplodon grayi Gray's beaked whale 55/0 100/16 MpeJCR1926 MpeLAM95654/U13141 USNM504259 83/7 69/0 USNM504260 TMMC-C75 96/12 USNM504853 LAM088901 Mesoplodon peruvianus lesser beaked whale Mesoplodon perrini sp. n. Perrin's beaked whale MstSW10402/SW4962 100/15 MstSW9491/SW6481 Mesoplodon stejnegeri Stejneger's beaked whale MheNZ02/TAS 98/16 MheNMNZ2173/SAM16387 Mesoplodon hectori Hector's beaked whale Berardius bairdii Baird's beaked whale nucleotide divergence per lineage (GTR correction) 14% Figure 2. 7% 0% Treelength=289 Consistency index (CI)=0.653 Retention index (RI)=0.789 Phylogenetic relationships among Mesoplodon perrini, M. hectori, and sub- set of related Mesoplodon beaked whales, based on maximum likelihood (ML) analyses, using combined mitochondrial DNA control region and cytochrome b sequences (821 bp). Numbers adjacent to internal nodes are ML bootstrap values ≥50%/Bremer sup- port indices. The figures adjacent to the branch termini show the diagnostic size, shape, and position of the teeth in the lower jaw of the adult male of each species. The circle indicates the position of the diminutive tooth in M. peruvianus. The arrows draw attention to the morphological similarity between M. perrini and M. hectori. [PAGE BREAK] DALEBOUT ET AL. NEW SPECIES OF BEAKED WHALE 587 (3) Sub-terminal narrow teeth, up to 64 mm long, 47 mm wide and 12 mm broad, with smooth anterior margins and a 60°-70° terminal angle. (4) Narrow triangular subvertex. (5) Narrow, reverse V-shaped space between right and left nasals. (6) Narrow premaxillaries adjacent to the antorbital notches. (7) Small, but distinct basirostral groove. (8) Antorbital notches and prominences formed by the maxilla. (9) Margins of the posteromedial portion of the maxillaries angled sharply laterally. Osteology and Dentition MORPHOLOGICAL DESCRIPTION Cranial and mandibular measurements are shown in Table 2, 3, respectively. The premaxillary crest is relatively narrow and conservative in shape (Fig. 6), similar to that of M. hectori and M. peruvianus. The cranium of the holotype is not greatly inflated, adding to the narrow triangular appearance of the synvertex, which is constricted laterally at the confluence of the maxillaries with the supraocciptals. The margins of the posteromedial portion of the max- illaries (posterior to the synvertex) are angled sharply laterally, more so on the right side, and the sides of the cranium are steep. There are moderately formed maxillary crests above the orbits. The rostrum is relatively short (82%-93% zygomatic width [ZW]) for a Mesoplodon, and the mesorostral canal is fully ossified in the adult male. The mandibles have a short symphysis, the ventral profile of which is convex. The remaining ventral profile of the mandible is concave and then convex again near the posterior end. As in other species of this genus, the teeth are reduced to a single, laterally compressed pair in the mandible. The teeth are set close to the apex of the jaw, and are thus com- pletely anterior to the posterior end of the symphysis. In the adult male ho- lotype specimen, the anterior edge of the teeth is 23 mm from the tip of the lower jaw in life, with 33 mm of the tooth exposed above the gumline. The shape of the exposed portion of the tooth is a rough isosceles triangle, but with a smoothly convex anterior margin. The teeth have a sharp terminal angle of between 60° and 70°, and splay outwards from the perpendicular by an angle of approximately 15° (Fig. 7, 8e). The teeth of the adult female (USNM504260) are sharply triangular, and are not erupted. Vertebral counts, based on the four type specimens are as follows: cervicals 7 (1 and 2 fused), thoracic 9-10, lumbars 11-13, total 46-50. The phalangeal formula (based on two specimens) is I-2, II-6, III-7, IV-5, V-3 or 4. COMPARISONS WITH MESOPLODON HECTORI From the original misidentification of four of the five specimens of M. perrini known to date (Mead 1981, Mead and Baker 1987), it is clear that this species resembles M. hectori (Gray, 1871) morphologically. Further examination of the [PAGE BREAK] 588 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 10 20 30 40 50 60 TMMC-C75 USNM504259 USNM504260 ?????????? ?????????? ?????????? ?????????? ?????????? ??. USNM504853 ?????????? ?????????? ?????????? ?????????? ?????????? ??. LAM088901 ?????????? ?????????? ?????????? ?????????? ?????????? ??. MheNMNZ2173 GAAAAAGTCT TGTTATAGAA TCACCATAAC CTTACAGTAC TACGTCAGTA TTGAAAAAGA .CT. .A..G..... MpeLAM95654 70 80 ..C... .C. ..T. A.. 90 100 110 120 TMMC-C75 AATACCCTAC AGTACATTTA CTGTATTAAT CATACAGACA CATTCA-CTG AGCGCTAATA USNM504259 USNM504260 USNM504853 LAM088901 MheNMNZ2173 MpeLAM95 654 T.T. TMMC-C75 USNM504259 USNM504260 USNM504853 LAM088901 MheNMN22173 MpeLAM95 654 TMMC-C75 USNM504259 USNM504260 USNM504853 LAM088901 MheNMNZ2173 MpeLAM95 654 130 140 150 .A. ..CC..C..A GA.A. ..CC..C..A 160 170 180 TATAGCGTCT CTCTAAAAGT GTATGTATAT ATATACTATG TATAACTGTG CATTCATTTA 190 TC.G..C. 200 GT 210 220 230 240 TTTTCACTAC GGAGAGTTAA AGCTCGTAAT TAATTTTTTT TATTTTACAT AAGTACATAA 250 260 270 280 290 300 TTTGCATTAT TCGTACATGT GCCCGTTCCA TTAGATCACG AGCTTAATCA CCATGCCGCG TMMC-C75 USNM504259 USNM504260 USNM504853 LAM088901 MheNMNZ2173 ..A MpеLAM95 654 ?? 310 320 3.30 340 350 360 TMMC-C75 TGAAACCAGC AACCCGCTTG GCAGGGATCC CTCTTCTCGC ACCGGGCCCA TCGATCGTGG USNM504259 USNM504260 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? USNM504853 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? LAM088901 MheNMNZ2173 MpeLAM95 654 370 380 390 400 410 420 TMMC-C75 GGGTAGCTAA TATTGCCTTT TATAAGACAT CTGGTTCTTA CTTCAGGACC ATCTTAACTT USNM504259 ?????? ?????????? ?????????? ?????????? ?????????? ?????????? USNM504260 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? USNM504853 ?????????? ?????????? ?????????? ?????????? ?????????? ?????????? LAM088901 MheNMNZ2173 MpeLAM95 654 ..T.. TMMC-075 430 AAAATCGCCC ACTC USNM504259 ?????????? ???? USNM504260 ?????????72 ???? USNM504853 LAM088901 ?????????? 7777 MheNMNZ2173 MpeLAM95 654 [PAGE BREAK] DALEBOUT ET AL. NEW SPECIES OF BEAKED WHALE 589 mandibles, teeth and skulls of M. perrini and M. hectori, including new material from the latter species held in Southern Hemisphere museum collections, show distinctive differences between the two species. These differences are docu- mented below. Mandibles and Teeth The mandibles of M. perrini have a short symphysis (19.3%-23.2% of man- dible length) compared to M. hectori (25.9% -33.8% of mandible length). The ventral lateral outline of the rami is convex over the length of the symphysis in M. perrini, whereas in M. hectori this outline is concave (Fig. 9). The entire ventral margin of the M. hectori mandible consists therefore of two concave areas, giving the rami a more slender appearance. This characteristic mandible shape in M. hectori was noted by Harmer (1924). In specimens of M. perrini, the teeth are situated slightly posterior (1-2 cm) to the tip of the mandibles. In M. hectori, this is only the case in juvenile and sub-adult specimens; the fully adult male teeth are situated at the very tip of the mandibles. In M. perrini, the alveoli are only slightly expanded to take the teeth, which reach 12 mm in breadth in adult males, giving the jaw an smoothly attenuating tip. In M. hectori, the adult male teeth are broader (<17.5 mm) and the alveoli are consequently expanded, such that the tip of the jaw is swollen. There are small differences in the teeth of these species. The teeth in M. perrini have, in profile, a smoothly convex anterior margin, whereas those of M. hectori have a margin with three flattish areas between the denticle and root (Fig. 10). In subadult specimens of M. hectori, these "flats" are more defined and appear as steps in the margin. In situ, the teeth of M. perrini are more erect, with a shorter posterior margin that those of M. hectori. The angle formed by the denticle is 60°-65° in M. perrini and 85°-90° in M. hectori. The female teeth of M. perrini are thin and sharply triangular with straight or slightly concave margins, while those of M. hectori have the same flat areas in the anterior margin as do the males of that species. Cranial Morphology The main difference in cranial morphology between these two species is in the shape of the neurocranium and synvertex. In M. perrini, the synvertex Figure 3. Aligned sequences for Mesoplodon perrini over 434 bp of mitochondrial DNA control region. Identity to reference sequence, TMMC-C75, indicated by dots. All five specimens of M. perrini share same haplotype over this fragment. Sequences from M. hectori MheNMNZ2173 (underlined) and M. peruvianus MpeLAM95654 in- cluded for comparison. Position 1 of alignment corresponds to position 15891 of fin whale, Balaenoptera physalus, mtDNA genome (Arnason et al. 1991). Diagnostic nu- cleotide position distinguishing M. perrini from all 20 previously described species of beaked whales at this locus highlighted in gray [PAGE BREAK] 590 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 10 20 30 40 50 ATG ATC AAC ATT CGA AAA ACA CAC CCA TTA ATA AAA ATT ATC AAC AAC GCA TTT TMMC-C75 USNM504259 ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? USNM504260 777 277 277 727 222 ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? USNM504853 777 777 777 777 777 777 777 277 227 222 227 277 ??? ??? ??? ??? ??? ??? LAM088901 MheSAM16387 .C... Mpe-013141 ....C. 60 ..T... C.. C.G 70 80 ..C ..C G.T..T..T T 90 100 ..C TMMC-C75 ATT GAT CTT CCC ACT CCA TCA AAC ATC TCC TCA TGA TGA AAC TTT GGT TCA TTA USNM504259 ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??. USNM504260 ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??.... USNM504853 777 27? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? LAM088901 777 777 ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??? ??. MheSAM16387 Mpe-U13141 ..C 110 ...T.C..G... T ..T ..T 120 130 140 150 160 TMMC-C75 CTC GGT CTC TGC CTA ATC ATA CAA ATT CTT ACA GGC TTA TTC CTA GCA ATA CAC USNM504259 USNM504260 USNM504853 LAM088901 MheSAM16387 Mpe-U13141 ??? ?? ??? ??? ?.. TMMC-C75 USNM504259 USNM504260 USNM504853 LAM088901 MheSAM16387 Mpe-U13141 TMMC-C75 USNM504259 USNM504260 USNM504853 LAM088901 MheSAM16387 Mpe-U13141 TMMC-C75 USNM504259 USNM504260 USNM504853 LAM088901 Mhe SAM16387 Mpe-U13141 170 180 190 200 210 TAC ACA CCA GAC ACT ACA ATA GCC TTT TCA TCA GTT GCA CAC ATC TGC CGA GAC 220 :::: 230 240 250 ..T 260 270 GTT AAC TAT GGT TGA ATT ATC CGA TAC CTA CAT GCA AAT GGA GCT TCC ATA TTT 280 290 300 310 320 TTT ATC TGC CTT TAC GCA CAC ATT GGA CGC GGT CTA TAT TAC GGC TCT TAT ATT 330 340 350 360 370 TTT CAA AAA ACA TGA AAT ATT GGA GTA ATT TTA CTC TTC ATA GTT ATA GCT ACT TMMC-C75 USNM504259 USNM504260 USNM504853 LAM088901 G.. MheSAM16387 ..C Mpe-U13141 380 TMMC-C75 GCA TTT USNM504259 USNM504260 USNM504853 LAM088901 MheSAM16387 Mpe-U13141 Figure 4. Aligned sequences for Mesoplodon perrini over 384 bp of mitochondrial DNA cytochrome b. Identity to reference sequence, TMMC-C75, indicated by dots. Two variable sites, defining three unique haplotypes, were found among the five spec- imens of M. perrini. Sequences from M. hectori MheSAM16387 (underlined) and M. peruvianus Mpe-U13141 are included for comparison. Position 1 of alignment corre- sponds to position 14613 of fin whale mtDNA genome (Arnason et al. 1991). Diag- [PAGE BREAK] DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE 591 narrows upwards, and the upper part of the skull is triangular in frontal view. In M. hectori, the synvertex is flatter, giving the upper part of the skull a rounded box-like shape. In M. perrini, the maxillaries below and behind the synvertex on each side of the neurocranium are steep-sided and not greatly inflated, whereas in M. hectori, the neurocranium is inflated and the maxillaries are prominent in frontal view on each side of the synvertex. Also, in M. perrini the margins of the posteromedial portion of the maxillaries at the uppermost level are sharply angled laterally on each side (90° from the longitudinal me- dian line on the right side, and 110° on the left). In M. hectori, these angles are usually gentle (130°-140°) although one specimen of M. hectori from South Africa (PEM 1511/15; Ross 1970) was found to have right side maxillary with a 90° angle. Therefore, this feature may not be a reliable distinguishing char- acter for M. perrini. Associated with the synvertex is the space between the right and left nasals. This is narrow and reverse V-shaped in M. perrini, but wide with parallel sides in M. hectori. The width of this space can be expressed as the least distance between the anterior prominences of the synvertex, which in M. perrini is 2.1%-6.7% ZW, and in M. hectori, 8.3%-18.5%. The premaxillaries of M. perrini are noticeably narrower than M. hectori. At the position of the antorbital notches, the width of M. perrini is 34.5%-35.4% (relative to the distance between the apices of the antorbital notches) compared to 43.3%-53.3% for M. hectori. At the position of the superior nares, the width of M. perrini is 35.2% -37.2% (relative to ZW) compared to 46.0%- 49.0% for M. hectori. There are a number of other, readily visible but less quantifiable features which separate the skulls of these two species, the most significant of which are: (1) In adult males, the mesorostral groove is fully ossified in M. perrini, but unossified in M. hectori (unusual in Mesoplodon, but the largest mature crania (695 mm and 677 mm CBL) of both sexes of M. hectori show little ossification). (2) M. perrini has a basirostral groove extending anterior to the maxillary prominences, while in M. hectori this groove is very short in adults and does not extend forward of the prominences, (3) In M. perrini, the antorbital notch is formed by the maxillary overlaying the jugal, and the antorbital prominence is formed by the maxillary; in M. hectori the antorbital notch is formed by the jugal, and the antorbital promi- nence is formed by the lacrimal. (4) Moderately formed maxillary crests are present above the orbit in M. perrini, but not in M. hectori. (5) In lateral view, the rostrum of M. perrini is deep through to mid-length nostic nucleotide position distinguishing M. perrini from all 20 previously described species of beaked whales at this locus highlighted in gray. [PAGE BREAK] frequency b 80 70 70 60 60 50 50 40 30 20 20 10 frequency 592 a 90 80 88 70 70 60 50 40 30 20 10 MARINE MAMMAL SCIENCE, VOL 18, NO. 3, 2002 Mesoplodon perrini sp. n. versus all other beaked whale species (average, 8.55%) intra-specific inter-specific pairwise sequence divergence (GTR-corrected) mtDNA control region low-level divergence between Arnoux's & Baird's beaked whales, B. arnuxii & B. bairdii 8%. %6 10%. 11%. 12% 13%- 14%. 15% intra-specific inter-specific pairwise sequence divergence (GTR-corrected) mtDNA cytochrome b %91 17% %81 Mesoplodon perrini sp. n. versus all other beaked whale species (average, 15.24%) 19% 20% 21% [PAGE BREAK] DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE 593 and thereafter tapers to the tip. In M. hectori, the outline is smoothly concave from the pterygoids to the rostrum tip. Although Harmer (1924) suggested that the structure of the thin part of the mesethmoid which forms the nasal septum was diagnostic for M. hectori, our observations suggest that this is a variable character. No significant dif- ferences were found in this feature among the specimens of M. hectori and M. perrini examined. The presence of a dorsal sinus also varies between individuals, and in both species, the septum rises above the level of the premaxillae in the region of the anterior nares. PREVIOUS ATTRIBUTIONS TO MESOPLODON HECTORI IN THE LITERATURE A number of publications, in addition to those already mentioned (Mead 1981, Mead and Baker 1987, Henshaw et al. 1997), have erroneously used data from specimens of M. perrini to represent M. hectori. In Mead (1984), reproductive data from specimens of M. perrini was used for M. hectori. Mead (1989) presented drawings of the cranium, mandible, and a tooth, as well as photographs of the external appearance of the holotype of M. perrini (USNM504853), as M. hectori. One of these photographs was used by Baker (1990, 1999). Some of these photographs also appeared in Reeves and Leath- erwood (1994). Jefferson et al. (1993) included some of the figures presented in Mead (1989). In Mead (1993), information on the stomach morphology of M. perrini was used for M. hectori. The artist's impression of M. hectori in Carwardine (1995) was based on photographs of the holotype of M. perrini. Messenger and McQuire (1998) used the mtDNA control region sequence from the calf, USNM504259 (Henshaw et al. 1997) to represent M. hectori in their phylogenetic analyses. Note that we have asked that this Genbank entry be corrected to reflect the true species identity of this specimen. It is possible that there are further publications that we are not aware of which have also made the mistake of using specimens of M. perrini to represent M. hectori. As no adult male specimens of M. bectori were known until relatively recently, images of the holotype of M. perrini have unfortunately been used widely to represent this species. External Appearance OTHER CHARACTERISTICS The overall body shape of M. perrini is typical of Mesoplodon beaked whales, with a relatively small head, a long thorax and abdomen, a deep peduncle, Figure 5. Frequency distribution of pairwise sequence divergence within and be- tween the 20 previously known species of beaked whales, based on: (a) 437 bp align- ment of mtDNA control region and (b) 384 bp alignment of mtDNA cytochrome b. Two representatives used per species where possible. GTR distance correction used to adjust for multiple substitutions. All intraspecific (open bars) and interspecific distances (shaded bars) are shown. [PAGE BREAK] 594 MARINE MAMMAL SCIENCE, VOL 18, NO. 3, 2002 Table 2. Cranial measurements for Mesoplodon perrini in mm. Methods of taking measurements follow Moore (1963) taken on right hand side where possible. No cranial measurements taken for USNM504259, as skull was crushed. Measurements currently not available from TMMC-C75. E, estimated length. Museum number Measurement number USNM504853 USNM504260 LAM088901 563 594 424 511 554 365 377 414 263 412 446 264 446 483 314 501 541 371 310 334 221 262 261 203 271 282 208 10 271 276 207 11 191 208 162 12 226 232 178 13 99 97 76 14 46 37 34 15 66 61 51 16 54 43 38 17 25 27 19 18 50 59E 35 19 20 21 24 25 26 27 28 29 30 31 33 34 35 36 37 38 39 222222222-23eneran 10 16 19 15 23 25 14 105 110 88 41 40 28 96 97 77 96 98 73 159 160 108 103 115 67 61 62 45 32 36 18 31 42 25 42 39 32 32 29 23 48 40 31 259 267 186 168 185 137 74 69 64 41 43 38 81 88 78 311 346 246 40 284 317 216 41 236 259 179 42 416 452 43 449 44 90 80 [PAGE BREAK] DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE Table 2. Continued. Museum number USNM LAM 504260 088901 70 54 30 male/calf Measurement USNM number 504853 45 64 46 595 sex/age class male/adult 205 female/adult greatest Definitions of cranial measurements (numbers in parentheses refer to Moore 1963): 1 = condylobasal length (1), 2 = tip rostrum to posterior extension maxillary plate (7); 3 tip rostrum to anterior margin superior nares (8); 4 tip rostrum to anterior point maxillary crest (9); 5 tip rostrum to posterior extension premaxilla on lateral tip of right premaxillary crest (11); 6 = tip rostrum to posterior extension temporal fossa (10); 7 tip rostrum to apices of antorbital notches (2); 8 breadth skull across orbital centres (19); 9 = breadth skull across postorbital process frontals (17); 10 = breadth skull across zygomatic processes squamosals (18), 11 = least breadth skull across posterior margins temporal fossae (20); 12 = greatest breadth skull across ex- occipitals (25); 13 = greatest span occipital condyles (21); 14 greatest width of an occipital condyle (22); 15 greatest length of an occipital condyle (23); 16 = breadth foramen magnum (24); 17 = greatest length of right nasal on vertex (15); 18 length nasal suture (16); 19 = extension right premaxilla posterior to right nasal on vertex (28); 20 greatest breadth nasals on vertex (26), 21 = least distance between anterior prominences of the synvertex (27); 22 = greatest span premaxillary crests (29); 23 = greatest transverse width of superior nares (37); 24 = least width premaxillae where narrow opposite superior nares (30); 25 greatest width premaxillae anterior to position of previous (31); 26 = width rostrum in apices of antorbital notches (33); 27 = width rostrum in apices of prominential notches (34); 28 = least distance be- tween main maxillary foramina (41); 29 = least distance between premaxillary foramina (42); 30 = distance posterior margin of left maxillary foramina to anterior margin maxillary prominence (43); 31 = width rostrum at mid-length of rostrum (35); 32 = width premaxillae at mid-length of rostrum (32); 33 depth rostrum at mid-length rostrum (36), 34 height of skull (39); 35 = external cranial height; 36 = greatest length of temporal fossa (13); 37 = width of temporal fossa (40); 38 = length of orbit taken from mid-point of frontals (14); 39 tip rostrum to posterior extension of maxilla between pterygoids (6); 40 = tip rostrum to anterior extension of pterygoid sinus (12); 41 = tip rostrum to most anterior extension of pterygoids (5); 42 rostrum to posterior margin of pterygoid mid-line (3); 43 = tip rostrum to posterior extension of wing of pterygoid (4); 44 length of vomer visible at surface of palate (44); 45 width between pterygoid notches (38); 46 = amount added to rostrum because of breakage (45). tip and short tail (Fig. 8a-e). The rostrum is relatively short compared to all other species in the genus, except M. hectori and M. peruvianus. In calves, the rostrum appears to be shorter and stubbier than in adults. The blowhole is broad and crescent-shaped, with anterior-pointing tips. The melon forms a small bulge, and the mouthline is straight. Throat grooves are present. External measure- ments are shown in Table 4. The adult male (USNM504853) was dark gray dorsally grading to white ventrally. The ventral side of the tail flukes was pale gray with converging striations and there was a white patch around the umbilicus. The adult female [PAGE BREAK] 596 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 Table 3. Mandibular measurements for Mesoplodon perrini. Measurements follow methods of Moore (1963), taken on right hand side where possible. No measurements available for USNM504259 as mandible has been lost. Measurements currently not available from TMMC-C75. E, estimated length. Museum number Measurement USNM USNM number 504853 504260 LAM 088901 47 516 486 356 48 387 379 289 49 455 402 314 50 120 108 69 51 97 52 37E 21 53 35 37 25 54 47 25 55 109 56 16 19 57 52 64 58 47 59 12 60 n/a 33 n/a 61 9.9 g³ sex/age class male/adult 37.0 gh female/adult male/calf right tooth, dry weight. b right tooth, wet weight. Definitions of measurements (numbers in parentheses refer to Moore 1963): 47 = mandibular length (1); 48 = length from posterior extension of symphysis to condyles (6); 49 = length posterior margin of alveolus to condyles (7); 50 = greatest length of symphysis (2); 51 greatest height of mandible at coronoid processes (3); 52 = outside height of mandible at midlength of alveolus (4); 53 = inside height of mandible at midlength of alveolus (5); 54 = length of alveolus (8); 55 = width of alveolus (9); 56 tip of mandible to alveolus (10); 57 = greatest tooth length (11); 58 greatest tooth width (12); 59 greatest tooth breadth (13); 60 = height of crown of tooth; 61 = tooth weight. (USNM504260) was too decomposed to allow information on color pattern to be collected. Calves (USNM504259, LAM088901 and TMMC-C75) are light to dark gray dorsally and white ventrally (Fig. 8a-d). The lower jaw and throat regions are white. A dark gray region extends from the corner of the mouth and encompasses the eye and the rostrum, forming an extended mask. The flippers are medium to dark gray dorsally and white ventrally. There is a lighter-colored patch on the anterodistal portion. The flukes are dark gray dorsally and medium to light gray ventrally. The ventral surface includes a pattern of white striations that converge posteromedially. Ontogeny and Reproduction Both adult specimens (USNM504260, female, and USNM504853, male) were considered physically mature based on the fusion of the thoracic epiphyses [PAGE BREAK] a b DALEBOUT ET AL. NEW SPECIES OF BEAKED WHALE 597 Figure 6. Skull photographs, USNM504853, holotype: (a) lateral view, (b) dorsal view. Scale pattern in (b) = 5 cm, divided into 50-mm squares, and pertains to both figures. to the centra and the disappearance of the epiphyseal suture (Mead 1981). While the mandibular rami of the male were fused at the symphysis, those of the female were not. The teeth of the male were fully erupted (Fig. 7), and the mesorostral canal completely filled, suggesting that it was sexually mature. The testis weights were; 84.5 g (L), and 115.5 g (R). The adult female ap- peared to have been dead for about two weeks when discovered (Mead 1981), and was too decomposed for the reproductive organs to be recognizable. It is likely that this animal was the mother of the calf (USNM504259), which had been found in approximately the same location a week earlier (Table 1). Both [PAGE BREAK] 598 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 a b [PAGE BREAK] DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE 599 animals share the same haplotype at the mtDNA control region and cyto- chrome b (Fig. 3, 4), as expected for a cow-calf pair. Although there has been little opportunity to date for calibration and standardization in the aging of Mesoplodon species, thin tooth sections were prepared from the right tooth of both adult specimens, and the cemental growth layer groups (GLG's) counted, giving an approximate age of nine years each, although the male appeared the more mature of the two from cranial features (Mead 1981). The 245-cm calf (LAM088901) had a squid eye lens in its stomach, which suggests that it had been weaned. There are no data on stomach contents for the other two calves (USNM504259 and TMMC-C75), but the fimbriated edge on the tongue of the former animal suggests that it was still suckling (Fig. 8d). The Monterey calf (TMMC-C75) had small, immature testes, which were elliptical in shape and weighed approximately 1.5 g each. NATURAL HISTORY Food Habits Only two of the animals had stomach contents. The stomach of a third (TMMC-C75) was collected, but is currently not available for examination. A squid eye lens, not identifiable to species, was found in the stomach of the second calf (LAM088901). Two lower beaks of the squid, Octopoteuthis deletron, and a fragment of an unidentifiable vertebrate were found in the stomach of the adult female (USNM504260; Mead 1981). We assume that like many other beaked whales, this species mainly eats pelagic squid. Behavior The adult male (USNM504853) bore a number of white, linear scars on its postcranial flanks which were probably inflicted by the teeth of other males of the species. However, it is noted that the scars on the adult male appear to have been made with a single tooth, rather than with two teeth simultaneously, as might be expected in species with apical teeth (e.g., Heyning 1984). Parasites Three soft-stalked barnacles, Conchoderma auritum, were found on the teeth of the adult male (USNM504853), and a number of cysts of the cestode, Phyllobothrium sp., were found encased in the blubber. Several oval scars (ap- proximately 3 x 5 cm) were found on the flanks. The Monterey calf (TMMC- C75) bore three similar scars, in various stages of healing. Such scars are likely due to cookie-cutter shark attacks (Isistius spp.; Jones 1971). This calf was Figure 7. Teeth of adult male (USNM504853) in situ: (a) oblique lateral view; (b) anterior view. Scale bars = 30 mm. Photograph credit: J. G. Mead. [PAGE BREAK] 600 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 b d e Figure 8. External photographs. (a) left lateral view of calf (TMMC-C75), scale bar = 30 cm; (b) ventral view of calf (TMMC-C75), scale bar 30 cm; (c) right lateral view of calf (TMMC-C75), scale bar = 30 cm; (d) right lateral view of head of calf = 20 cm; (e) left lateral view of head of adult male (USNM504259), scale bar (USNM504853), scale bar = 24 cm. Note that striped pattern on right lateral side of calf (TMMC-C75) is the imprint caused by bed of utility truck on which animal was lying. Photograph credits: (a-c) M. Haulena; (d, e) G. Carsten and J. G. Mead. [PAGE BREAK] C DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE 601 M. hectori M. hectori 50mm Figure 9. M. perrini Lateral view of mandibular ramus of Mesoplodon hectori (a, NMNZ2173 adult male; b, NMNZ614 juvenile male) and M. perrini (c, USNM504853 adult male). M. perrini 40mm M. hectori Figure 10. Adult male tooth of Mesoplodon perrini (USNM504853) and M. hectori (NMNZ2173). Right tooth of each species is shown, with anterior margin facing left. [PAGE BREAK] 602 External measurements for Mesoplodon perrini. Due to extent of decomposition, few measurements were available from adult female Table 4. (USNM504260). Museum or field number: MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 USNM504853 Male/adult USNM504260 Female/adult USNM504259 Male/calf LAM088901 Male/calf TMMC-C75 Male/calf Sex/age class: Measurement: cm cm cm cm cm Total length 390.0 100.0 443.0 100.0 210.0 100.0 245.0 100.0 223.5 100.0 Snout to center of blowhole 35.1 9.0 22.7 10.8 27.0 11.0 18.5 8.3 Snout to center of eye Snout to angle of mouth Snout to ear Snout to anterior insertion of flipper Snout to center of umbilicus Snout to genital slit (center) Snout to anus 42.5 10.9 25.3 12.0 28.5 11.6 25.5 11.4 24.0 6.2 19.5 9.3 15.2 6.2 14.0 6.3 50.2 12.9 29.6 14.1 33.7 13.8 72.0 18.5 48.4 23.0 51.7 21.1 47.0 21.0 150.0 38.5 123.4 50.4 105.5 47.2 254.0 65.1 140.0 66.7 162.0 66.1 142.0 63.5 285.0 73.1 159.0 75.7 184.8 75.4 160.1 71.6 Snout to tip of dorsal fin 240.0 61.5 274.0 61.9 140.0 66.7 162.9 66.5 146.0 65.3 Girth at anus Girth at eye Girth at axilla Maximum girth Girth midway anus to fluke notch 120.0 30.8 84.5 34.5 72.5 32.4 196.0 50.3 127.0 51.8 107.0 47.9 230.0 59.0 119.5 53.5 130.0 33.3 36.7 70.0 31.3 66.0 16.9 48.0 21.5 Height at same place as above 30.6 7.8 24.0 10.7 Thickness at same place as above 9.0 2.3 9.5 4.3 Projection lower/upper jaw 1.6 0.4 0.6 0.3 0.6 0.2 2.0 0.9 Length of eye opening 3.2 4.0 1.8 0.8 Center of eye to ear 9.0 2.3 5.4 2.6 7.0 2.9 Center of eye to angle of mouth 18.8 4.8 6.0 2.9 17.1 7.0 12.5 5.6 Blowhole width 9.9 2.5 4.8 2.3 5.8 2.4 5.7 2.6 Length of throat grooves 20.0 5.1 9.7 4.6 19.7 8.0 14.0 6.3 693 [PAGE BREAK] Museum or field number: USNM504853 Table 4. Continued. USNM504260 USNM504259 Male/adult Female/adult Male/calf LAM088901 Male/calf TMMC-C75 Male/calf Sex/age class: Measurement: cm cm cm cm cm Flipper length, anterior 46.0 11.8 40.0 9.0 28.0 13.3 31.5 12.9 30.5 13.6 Flipper length, posterior Flipper width, maximum Fluke width Fluke depth 29.0 7.4 32.0 7.2 19.0 9.0 20.1 8.2 24.0 10.7 13.2 3.4 7.6 3.6 9.0 3.7 8.5 3.8 103.0 26.4 89.0 20.1 46.7 22.2 52.0 21.2 58.0 26.0 32.0 8.2 18.0 8.6 20.8 8.5 19.0 8.5 Depth of fluke notch NA 1.9 0.9 NA 1.2 0.5 Dorsal fin height 19.0 4.9 10.2 4.9 13.6 5.6 9.0 4.0 Length dorsal fin base 50.0 12.8 23.0 11.0 18.0 8.1 603 [PAGE BREAK] 604 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 severely emaciated, with minimal body fat and atrophied muscles. No parasites were found in gross necropsy, but a histological examination of abscesses in the periumbilical region revealed that they contained degenerating cestodes (likely Phyllobothrium spp., possibly Phyllobothrium delphini) and other foreign material. Structures resembling parasitic granulomas were found within the gastrointestinal tract and one of the lymph nodes (necropsy performed by F. Gulland, University of California at Davis; details courtesy of M. Haulena). Distribution This species is known from five specimens found beachcast along the Cal- ifornian coast between Torrey Pines State Reserve, just north of San Diego (32°55'N, 117°15'W) and Fisherman's Wharf, Monterey (36°37'N, 121°55'W). Although this stranding pattern is suggestive of an eastern North Pacific distribution, there are too few records to date to draw any bounds on this. Little can be concluded from the presence of cookie-cutter shark scars on the Monterey calf. Isistius spp. are limited in their northern distribution, at least in surface waters (Nakano and Tabuchi 1990), but the occurrence of such scars on cetaceans is not (Jones 1971). This suggests either that these cetaceans are migratory and pass through the territory of Isistius spp., or that the dis- tribution of Isistius spp. extends farther north in deeper waters and they attack cetaceans when they dive. Given the habitat preferences of other ziphiids, we assume that M. perrini is found primarily in oceanic waters, over 1,000 m in depth. DISCUSSION According to the International Code of Zoological Nomenclature (ICZN), formal classification of M. perrini ideally requires genetic validation of the holotype of M. hectori, as four of the five known specimens of M. perrini were described previously as this species (Mead 1981). The holotype of M. hectori is held by the British Museum (BM(NH) 1677/76.2.16.3), and consists of the skull, mandible, scapulae, hyoids, cervical vertebrae and flippers of a juvenile male collected in Titahi Bay, Wellington, New Zealand, in the 19th century (Flower 1878; Gray 1871). We have attempted, but not been successful in extracting and amplifying native DNA from the M. hectori holotype. This may be due to the coat of varnish on this specimen, the acidic components of which can degrade the already low levels of endogenous DNA contained in skeletal material (Cooper 1994). In the absence of genetic data from the holotype of M. hectori, validated specimens from New Zealand and Australia have been used to represent this M. hectori in these analyses. Given the increasing use of genetic information as a universal character in species identification, systematics, and biodiversity assessment (Bisby 2000, Wilson 2000), the designation of genetic voucher material should be consid- ered for all taxa. It has been suggested that mtDNA typing of holotype spec- imens should become part of standard museum protocol (Dalebout 2002), but [PAGE BREAK] DALEBOUT ET AL. NEW SPECIES OF BEAKED WHALE 605 it is recognised both that: (1) not all specimens will retain sufficient native DNA for analysis due to a combination of age, the nature of the material available (e.g., bone, tooth, pelts, feathers), and museum preparation and stor- age methods and (2) not all specimens will be amenable to current DNA extraction techniques (e.g., Wayne et al. 1999). In such cases, genetic material should be obtained from other validated specimens of the species in question (e.g., Dizon et al. 2000). Genetic voucher material, in the form of DNA se- quences from a suite of loci, could be archived both at the institution holding the holotype (or alternative validated specimen from which these data were derived), as well as in the international genetic database, Genbank. When describing new species, a genetic description should also be included wherever possible. A recent analysis of discovery trends has suggested that at least 40 species of large marine animals still remain to be described (Paxton 1998). If so, it is likely that several new cetacean species will be among them, including at least one new form of Bryde's whale, Balaenoptera edeni sp. (e.g., Baker et al. 1996, Yoshida and Kato 1999), and possibly further species of ziphiids. The current discovery represents the second new species of Mesoplodon discovered in the last decade. Both were dependent on the opportunistic collection of beachcast specimens and victims of fisheries bycatch (Reyes et al. 1991, this paper). Yet, there is no reason to assume that all cetacean species can be encountered in this way. Some may be distributed in areas far from shore or shore-bound currents, where human presence is still minimal. To ensure that such species do not go undocumented, we recommend that biopsy samples be collected wherever possible from animals encountered on sighting cruises. In addition to traditional morphological information, the collection of tissue sam- ples from stranded and incidentally caught animals should also become stan- dard procedure. ACKNOWLEDGMENTS We are extremely grateful to all those who gave us access to specimens used to establish the beaked whale DNA reference database that has been so valuable for these and other discoveries (see http://www.sbs. auckland.ac.nz/research-groups/ecology_and_evolution/ molecular ecol_evo_l_lab for a full list) For the work concerning M. perrini, we would especially like to thank the following people and institutions for collection and access to specimens, laboratory support, valuable discussion and other aid: field staff of the New Zealand Department of Conservation; A. E. Dizon, M. Henshaw, and K. Rob- ertson, NOAA SWFSC; J. E. Heyning, Los Angeles County Museum of Natural History; P. Jenkins, A. Warlow, R. Sabin, Natural History Museum (BMNH), London; S. J. O'Brien, N. Yuhki, Laboratory of Genomic Diversity, National Cancer Institute (also M. Culver, V. David, G. M. Lento, J. Martenson, B. Murphy, M. Raymond, J. Pecon-Slattery, and G. K. Pei); G. Carsten, J. M. Coe, J. R. Henderson, D. B. Holts, K. LeVeille, W. F. Perrin, W. A. Walker, M. Haulena, NOAA SWFSC and The Marine Mammal Centre; C. W. Potter, Smithsonian National Museum of Natural History; D. J. Saul, A. G. Rodrigo, and V. Ward, University of Auckland; K. Van Waerebeek, CEPEC, Peru. For access to specimens of M. bectori, we would like to thank C. Kemper, SAM, P. Sabine, TAM, and N. Prosser-Goodall, MAAMA. We would also like to thank our two reviewers, F. Cipriano and G. J. B. Ross, for their very useful comments on [PAGE BREAK] 606 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 the manuscript. MLD was supported by a William Georgetti Scholarship, a University of Auckland Doctoral Scholarship, and a Smithsonian Institution Short-Term Visitor's Fellowship. This project was supported by grants to C. S. Baker from the University of Auckland, the New Zealand Lottery Board, the New Zealand Marsden Fund, and the International Fund for Animal Welfare. LITERATURE CITED ARNASON, U., A. GULLBERG AND B. WIDEGREN. 1991. The complete nucleotide se- quence of the mitochondrial DNA of the fin whale, Balaenoptera physalus. Journal of Molecular Evolution 33:556-568. BAKER, A. N. 1990, 1999. Whales and dolphins of New Zealand and Australia: An identification guide. Victoria University Press, Wellington. 133 pp. BAKER, C. S., A. PERRY, J. L. BANNISTER, M. T. WEINRICH, R. B. ABERNETHY, J. CALAMBOKIDIS, J. LIEN, R. H. LAMBERTSEN, J. URBAN RAMIREZ, O. VASQUEZ, P. J. CLAPHAM, A. ALLING, S. J. O'BRIEN AND S. R. PALUMBI. 1993. Abundant mito- chondrial DNA variation and world-wide population structure in humpback whales. Proceedings of the National Academy of Sciences of the U.S.A 90:8239- 8243. BAKER, C. S., F. CIPRIANO AND S. R. PALUMBI. 1996. Molecular genetic identification of whale and dolphin products from commercial markets in Korea and Japan. Molecular Ecology 5:671-685. BISBY, F. A. 2000. The quiet revolution: Biodiversity informatics and the internet. [AD] Science 289:2309-2312. CARWARDINE, M. 1995. Whales, dolphins and porpoises. HarperCollins, London. COOPER, A. 1994. Dried samples: DNA from museum specimens. Pages 149-165 in B. L. Herman and S. Hummel, eds. Ancient DNA. Springer-Verlag, New York, NY. DALEBOUT, M. L. 2002. Species identity, genetic diversity and molecular systematic relationships among the Ziphiidae (beaked whales). Ph.D. thesis, School of Bio- logical Sciences, University of Auckland, Auckland, New Zealand (submitted for examination, October 2001). DALEBOUT, M. L., A. VAN HELDEN, K. VAN WAEREBEEK AND C. S. BAKER. 1998. Mo- lecular genetic identification of southern hemisphere beaked whales (Cetacea: Zi- phiidae). Molecular Ecology 7:687-694. DAVIS, J. I, AND K. C. NIXON. 1992. Populations, genetic variation and the delimi- tation of phylogenetic species. Systematic Biology 41:421-435. DIZON, A., C. S. BAKER, F. CIPRIANO, G. LENTO, P. PALSBØLL AND R. REEVES. 2000. Molecular genetic identification of whales, dolphins and porpoises: proceedings of a workshop on the forensic use of molecular techniques to identify wildlife prod- ucts in the marketplace. NOAA Technical Memorandum NMFS NOAA-TM- NMFS-SWFSC-286, La Jolla, CA. FARRIS, J. S., M. KALLERSJÖ, A. G. KLUGE AND C. BULT. 1995. Testing the significance of incongruence. Cladistics 10:315–319. FLOWER, W. H. 1878. A further contribution to the knowledge of the existing ziphioid whales. Genus Mesoplodon. Transactions of the Zoological Society, London. 10: 415-437. GRAY, J. E. 1871. Notes on the Berardius of New Zealand. Annals and Magazine of Natural History 4th series: 115-117. GRAY, J. E. 1874. Notes on Dr Hector's paper on the whales and dolphins of the New Zealand seas. Transactions of the New Zealand Institute VI:93-97. HARMER, S. F. 1924. Mesoplodon and other beaked whales. Proceedings of the Zoological Society XXXVI:36-587. HENSHAW, M. D., R. G. LEDUC, S. J. CHIVERS AND A. E. DIZON. 1997. Identification [PAGE BREAK] DALEBOUT ET AL: NEW SPECIES OF BEAKED WHALE 607 of beaked whales (family Ziphiidae) using mtDNA sequences. Marine Mammal Science 13:487-495. HEYNING, J. E. 1984. Functional morphology involved in intraspecific fighting of the [AD] beaked whale, Mesoplodon carlhubbsi. Canadian Journal of Zoology 62:1645-1654. Höss, M., AND S. PAABO. 1993. DNA purification from Pleistocene bones by a silica- [AD] based purification method. Nucleic Acids Research 21:3913-3914. JEFFERSON, T. A., S. LEATHERWOOD AND M. A. WEBBER. 1993. FAO species identifi- cation guide: Marine mammals of the world. United States Environment Pro- gramme; Food and Agriculture Organization of the United Nations (FAO), Rome. JONES, E. C. 1971. Isistius brasiliensis, a squaloid shark, the probable cause of crater wounds on fishes and cetaceans. Fishery Bulletin. 69:791-798. LENTO, G. M., M. HADDON, G. K. CHAMBERS AND C. S. BAKER. 1997. Genetic variation of Southern Hemisphere fur seals (Arctocephalus spp.): Investigation of population structure and species identity. Journal of Heredity 88:202-208. MATISOO-SMITH, E., J. S. ALLEN, T. N. LADEFOGED, R. M. ROBERTS AND D. M. LAMBERT. 1997. Ancient DNA from Polynesian rats: Extraction, amplification and sequence [AD] from single small bones. Electrophoresis 18:1534-1537. MEAD, J. G. 1981. First records of Mesoplodon hectori (Ziphiidae) from the northern hemisphere and a description of the adult male. Journal of Mammalogy 62:430- 432. MEAD, J. G. 1984. Survey of reproductive data for the beaked whales (Ziphiidae). Report of the International Whaling Commision (Special Issue 6):91-96. MEAD, J. G. 1989. Beaked whales of the genus Mesoplodon. Pages 349-430 in S. H. Ridgway and R. Harrison, eds. Handbook of marine mammals. Volume 4. Aca- demic Press, London. MEAD, J. G. 1993. The systematic importance of stomach anatomy in beaked whales. IBI Reports 4:75-86. MEAD, J. G., AND A. N. BAKER. 1987. Notes on the rare beaked whale, Mesoplodon hectori (Gray) Journal of the Royal Society of New Zealand 17:303–312. MESSENGER, S. L., AND J. A. McQUIRE. 1998. Morphology, molecules and the phylo- genetics of cetaceans. Systematic Biology 47:90-124. MOORE, J. C. 1963. Recognizing certain species of beaked whales of the Pacific Ocean. The American Midland Naturalist 70:396-428. NAKANO, H., AND M. TABUCHI. 1990. Occurrence of the cookie-cutter shark Isistius brasiliensis in surface waters of the North Pacific Ocean. Japanese Journal of Ich- thyology 37:60-63. PAXTON, C. G. M. 1998. A cumulative species description curve for large open water marine animals. Journal of the Marine Biological Association of the United King- [AD] dom 78:1389-1391. PICHLER, F. B., M. L. DALEBOUT AND C. S. BAKER. 2001. Non-destructive DNA ex- traction from sperm whale teeth and scrimshaw. Molecular Ecology Notes 1:106- 109. REEVES, R. R., AND S. LEATHERWOOD. 1994 Dolphins, porpoises and whales: 1994-98 Action plan for the conservation of cetaceans. International Union for Conservation of Nature and Natural Resource, Gland, Switzerland. REYES, J. C., J. G. MEAD AND K. VAN WAEREBEEK. 1991. A new species of beaked whale Mesoplodon peruvianus sp. n. (Cetacea: Ziphiidae) from Peru. Marine Mam- mal Science 7:1-24. REYES, J. C., K. VAN WAEREBEEK, J. C. CÁRDENAS AND J. L. YAÑEZ. 1995. Mesoplodon bahamondi sp. n. (Cetacea, Ziphiidae), a living beaked whale from the Juan Fer- nandez Archipelago, Chile. Boletim de Museo Nacional de Historia Natural, Chile 45:31-44. Ross, G. J. B. 1970. The occurrence of Hector's beaked whale, Mesoplodon hectori (Gray) in South African waters. Annals of the Cape Provincial Museum (Natural History) 8:195–204. [PAGE BREAK] 608 MARINE MAMMAL SCIENCE, VOL. 18, NO. 3, 2002 SAMBROOK, E., F. FRITSCH AND T. MANIATIS. 1989. Molecular cloning. Cold Spring Harbor Press, Cold Spring Harbor, NY. SANDERSON, M. J., AND M. J. DONOGHUE. 1989. Patterns of variation and levels of [AD] homoplasy. Evolution 43:1781-1795. SORENSEN, M. D. 1999. TreeRot, version 2. Boston University, Boston, MA. SWOFFORD, D. L. 1999. PAUP*: Phylogenetic analysis using parsimony* and other phylogenetic reconstruction methods. Sinauer Associates, Sunderland, MA. VAN HELDEN, A. L., A. N. BAKER, M. L. DALEBOUT, J. C. REYES, K. VAN WAEREBEEK AND C. S. BAKER. 2002. Resurrection of Mesoplodon traversii (Gray, 1874), senior synonym of M. bahamondi Reyes, Van Waerebeek, Cárdenas and Yañez, 1995 (Cetacea: Ziphiidae). Marine Mammal Science 18:609-621. WAYNE, R. K., A. MEYER, N. LEHMAN, B. VAN VALKENBURGH, P. W. KAT, T. K. FULLER, D. GIRMAN AND S. J. O'BRIEN. 1990. Large sequence divergence among mito- chondrial DNA genotypes within populations of eastern African black-backed jackals. Proceedings of the National Academy of Sciences of the United States of [AD] America 87:1772-1776. WAYNE, R. K., J. A. LEONARD AND A. COOPER. 1999. Full of sound and fury: The recent history of ancient DNA. Annual Review of Ecology and Systematics 30: 457-477. WILSON, E. O. 2000. A global biodiversity map. Science 289:2279. YOSHIDA, H., AND H. KATO. 1999 Phylogenetic relationships of Bryde's whales in the western North Pacific and adjacent waters inferred from mitochondrial DNA [AD] sequences. Marine Mammal Science 15:1269-1286. Received: 08 July 2001 Accepted: 14 January 2002 [PAGE BREAK] NOTE INTORNO ALLA DISTRIBUZIONE DELLA FAUNA VERTEBRATA NELL'OCEANO prese durante un viaggio intorno al Globo 1865-68 dai Professore ENRICO HILLYER GIGLIOLI MEMBRO DELLA SOCIETÀ GEOGRAFICA ITALIANA, DELLA ZOOLOGICA DI LONDRA DELL' ENTOMOLOGICA DI FIRENZE ECC. Con una Carta. Two- finned whalebone whate 75-76 FIRENZE, STABILIMENTO DI GIUSEPPE CIVELLI Via Panicale, 39. 1870. [PAGE BREAK] tri l'At- 806; mo- il più anco ogici gran- balc- assa- ora, il 10 mo pure pas- vista pinna iso- dei 1 go- janco oleva inna edere ferire com- uper- vidi. grossi pro- que- olari nbini come Balaenoptera. Sp.? Una grossa specie, la quale per quanto ho potuto giudicare vedendola imperfettamente per alcuni minuti, non sembrava diversa dalla B. mu- sculus dell'Atlantico, presentando come quella la pinna dorsale molto in- dietro; la parte esposta del corpo era di un bruno-nero, non fece vedere la tesla. Fu incontrata il 1° settembre 1866, allo sbocco del Golfo di Yeddo. Non meno di quattro balenottere e due balene sono state descritte c baltezzate in tutte le regole, da soli disegni cinesi e giapponesi, sono perciò specie più che dubbiosc. I Giapponesi danno ai grossi cetacci il nome generico di kugira. BALAENOPTERA? Sp. e gen. nov. Il dopo pranzo del 4 settembre 1867 nel Pacifico, (lat. 28' 34' S.; long, 88° 10' O. Gr.); mentre stavo pescando minuti organismi pelagici alla scala reale, fui interrotto da un grande getto d'acqua che venne quasi a spruzzarmi, e vidi emergere dal mare la schiena grigio-verdastra di una bella balenottera, sul culmine della quale si vedevano due pinne ben marcate, separate da uno spazio di circa due metri. Queste pinne erano grandi, c ben sviluppate relativamente alle dimensioni dell'animale, che poteva avere un 18 metri di lunghezza. La testa era proporzionata, il muso largo e ar- rotondato, le mascelle quasi uguali, quella inferiore però più sporgente; gli occhi, come in tutti i cetacei, erano piccoli, le aperture delle narici situate sulla sommità della testa, e poste l'una vicino all'altra, davano pas- saggio al getto d'acqua misto ad aria, che aveva richiamate la mia atten- zione; l'animale muoveva intanto la mascella inferiore, come se masticasse qualcosa. La coda era relativamente grande, come pure le pinne pettorali; e mentre l'animale si voltava sul fianco destro per tuffarsi sott'acqua, vidi essere il ventre di color bianco puro, la pelle del petto mi sembrò liscia e senza rughe. In questa occasione ebbi la rara opportunità di vedere chiaramente tutti i movimenti di quel grosso cetaceo, che nuotava di poco al di sotto della superficie, la coda è il vero motore, agisce precisamente come una potente clice, le pinne pettorali erano poco usate, ed allora aiu- tavano a cambiare la direzione, facendo da limoni supplementari. Questa balenottera girò intorno alla Magenta fregando i suoi fianchi contro il bordo, ed ebbi tutto l'agio di esaminarla bene per oltre un quarto l'ora, mentre si stava ammainando una lancia onde vederla più da vicino. Intorno ad essa erano molti pesci, non potci determinarne la specie. Alcuni dei nostri vollero cercare di darle la caccia, ma non avevamo i mezzi per questo, malgrado la buona volontà del nostromo, il quale andò a cercare la sua migliore fiocina, ed una sessantina di metri di buona func, ci voleva altro. Scendemmo nella lancia, ma l'animale spaventato dal rumore non tardo a sprofondarsi, e non comparve più. l'er quanto io mi sappia, questa è la prima balenoltera osservata con [PAGE BREAK] due pinne sul dorso, e mi credo in dovere di considerarla non solo sic- come specie nuova, ma come il tipo di un nuovo genere, pel quale pro- pongo il nome di AMPHIPTERA; questo genero sarebbe per ora caratterizzato dalla presenza di due pinne dorsali: augurando che presto qualche na- turalista sia abbastanza fortdnato per avere a sua disposizione il cadavere di uno di esse, onde dare al mondo scientifico una descrizione più com- pleta di un cetaceo così interessante. La specie tipo, e per ora unica, porterebbe il nome di AMPHIPTERA PACIFICA. Non è improbabile che si trovi pure sulle coste meridionali dell' Au- stralia, e su quelle della N. Zelanda. Come abbiamo detto, non si conoscono altre balenottere munite di due pinne mediane, è bensì vero che Rafinesque descrisse molti anni fa, un delfino con due pinne dorsali, l'Oxypterus Mongitori, dei mari della Sici- lia; ma d'allora in poi mai più visto. Ed i naturalisti Quoy e Gaimard che accompagnarono Freycinet nel suo viaggio di circumnavigazione (1817-20), descrissero, per averla veduta un momento a volo d'uccello, nel Pacifico tra le isole Sandwich e l'Australia, un'altra specie più grande, il Delphi- (nus (Oxypterus) rhinoceros, cosi chiamata per avere una pinna od una prominenza curva indietro posta sull'occipite; anch'essa non fu più rive- duta. BALAENA. I caratteri sono quasi tutti gli stessi di quelli che definiscono il genere precedente, le balene si distinguono però per la mancanza della pinna dorsale, per avere la testa relativamente molto più grossa, la bocca piu grande, a margini sinuosi, la mascella inferiore più grande e più profonda della superiore, i fanoni più sviluppati; il corpo più tozzo, e lo strato adiposo soltocutaneo assai più spesso. Sono le specie di questo genere che vengono pescate con maggior profitto. Balaena australis. Desmoul. Questa specie ha la testa più proporzionata che non la B. mysticetus, che rappresenta nei mari australi, ove ha una distribuzione molto estesa, trovandosi in certe stagioni nei paraggi del Capo di Buona Speranza, in- torno alla Nuova Zelanda, alla terra di Kerguelen, all'Australia meridio- nale, ed alle terre più meridionali dell' America. È stata in questi ultimi anni oggetto di una caccia assai attiva, resa facile per l'abitudine che aveva di avvicinare le coste all'epoca della ri- produzione; da Hobart Town (Tasmania) partivono molti bastimenti per quella caccia; ora, il numero di questi cetacei è assai diminuito ina è tuttora molto più considerevole che non quello della balena vera, nei mari artici. Dal 1830 al 1831 non meno di 91 bastimenti inglesi, montati da 2050 marinai, e rappresentando una portata di 30,083 tonnellate, pescavano la balena nei mari del Sud, mentre dal 1840 al 1843 non se ne contavano [PAGE BREAK] THE BOOK OF WHALES [PAGE BREAK] A.570 WRITTEN AND ILLUSTRATED BY RICHARD ELLIS ALFRED A KNOPF NEW YORK 1980 USEF ROGEN QUE U [PAGE BREAK] said that calling an animal a whale does not necessarily make it one. Cetaceans are found in all the oceans of the world, from deep within the polar ice to the tropics. There are freshwater dolphins, restricted to certain rivers; and one species, the whitefin dolphin, is found only in Tung Ting Lake and its adjacent waters in China. The killer whale is probably the most widely distributed of all cetaceans, found in polar, temperate, and tropical oceans all over the world. Many species are migratory, moving from one area to another in clearly defined routes, whereas in other species the movements and distribution are poorly known. In recent years certain species have turned up in unexpected locations; for example, Fraser's dolphin, Lagenodelphis hosei, which was previously known only from a skeleton found on a beach in Borneo, has now appeared in the eastern tropical Pacific, off Taiwan, off Australia, and in the Caribbean. There is even the possibility of still-undiscovered cetaceans. Some of the beaked whales are known only from single skeletons, and there may be Mesoplodons whose remains have not yet washed ashore and been discovered by knowledgeable cetologists. On a re- mote beach in Tierra del Fuego, R. N. P. Goodall (1978) discovered the collected remains of some four- teen species of cetaceans and added substantially to our knowledge of the range, habits, and osteology of many of them. In 1964, during a whale-marking and observation cruise off the coast of Chile, a number of cetologists saw and photographed many of the ex- pected species, including blue, fin, sei, sperm, and smaller cetaceans. (As whalers will attest, the west coast of South America is a particularly productive area for their efforts.) On November 24 and 27, how- ever, "there were eight sightings . of small whales which we could not identify" (Clarke et al. 1978). The investigators chased and photographed these odon- tocete, single-blowhole, beakless whales, which had light-colored heads and dark bodies, were in a school of fifteen to twenty animals, and were about 20 feet (6.0 meters) long. (Only Risso's dolphin, Grampus gri- seus, fits this general description, but it is not known to exceed a length of 13 feet.) To demonstrate the exciting state of basic cetological research, I quote the observer's concluding paragraph in full: We can only conclude that these whales seen off Chile in 1964 may be a new species, possibly of the genus Pseudorca or Grampus. It is clearly important to obtain specimens as soon as opportunity affords. The animal agrees well with the "undescribed whale" reported and figured by Wilson (1905, P. 472; 1907, p. 4 and Whales Plate I) as 20-30 feet long, black above but with some white around the mouth or chin, and characterized by a high dorsal fin "erect, pointed and sabre-shaped"; several were seen in the Ross Sea during the [AD] British National Antarctic Expedition (1901-1904), but they have never been reported since. way: In 1967 a sport-fishing boat captain saw eighteen to twenty small cetaceans near the island of Tobago in the northeastern Caribbean. He described them in "The heads back to the dorsal fin and abdo- this mens were white, and the backs brownish gray. On the larger ones, the dorsal fin was close to ca. 0.6m (2 feet) high and hooked slightly backward" (Erdman et al. 1973). These whales, which were about 16 to 18 feet (4.9 to 5.5 meters) long, might very well be the same kind of mysterious animal observed by Clarke off Chile. gray Some species exist in uncountable numbers, roam- ing the open oceans far offshore, but others, closer to human habitation, shipping, and industry, are all too easily counted-and killed. The California whale, which was known in historical times from the western Pacific and the eastern Atlantic, exists now as a relict population of some 15,000 animals that pass close to the shoreline of western North America. The Indus River susu, Plantanista minor, another primitive spe- cies, is probably the most endangered of all cetaceans, having been reduced to some five hundred-odd ani- mals in the Indus River while hunting continues. The cetaceans are perhaps the most highly special- ized of all mammals. They are more or less fishlike in shape and, like the fishes, have a caudal fin used for propulsion and pectoral fins used for steering. The caudal fin of a cetacean, commonly known as the flukes, functions in an up-and-down movement, whereas the fishes, whose tails lie in the vertical plane, use a side-to-side motion. (In defining the whale, Mel- ville wrote that it "is a spouting fish with a horizontal tail.") Most cetaceans have a dorsal fin, which is be- lieved to help stabilize the animal in the water. Neither the caudal fin nor the dorsal fin has any bony internal support structure, but both are composed solely of connective tissue. Although some species have a num- ber of hairs on the head, for the most part the mam- malian hair in cetaceans has been replaced in function by a layer of fat or blubber, which serves to insulate the animal against the cold of its warmth-absorbing habitat. Like all other mammals, whales and dolphins give birth to live young and nurse them by means of




